Quá trình thụ tinh, làm tổ của phôi và những thay đổi đầu tiên trong thai kỳ
Trungtamthuoc.com - Khi một tinh trùng kết hợp với trứng tạo ra hợp tử, di chuyển và làm tổ tại niêm mạc tử cung, hình thành nên phôi thai, mở đầu cho sự phát triển của thai nhi. Quá trình thụ tinh, làm tổ và phát triển phôi được gọi là giai đoạn tiền thai.
1 SINH LÝ THỤ TINH
1.1 Phản ứng cực đầu
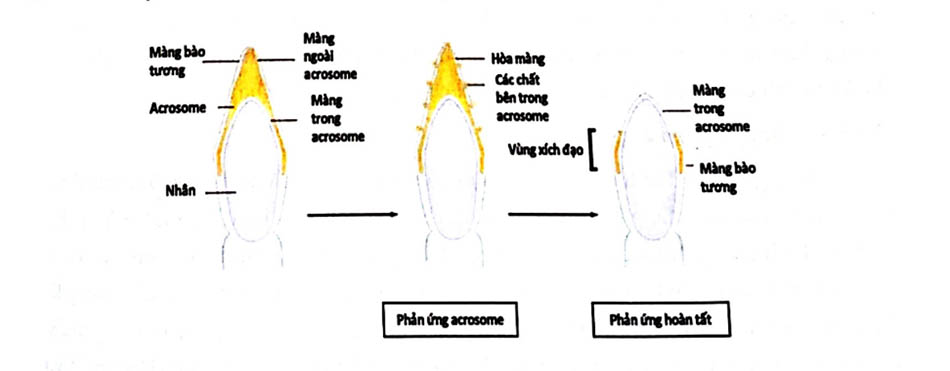
Phản ứng cực đầu là tình trạng thể cực đầu được “kích hoạt” hay bị phá vỡ để giải phóng các chất giúp tinh trùng xâm nhập vào noãn, là điều kiện tiên quyết để thực hiện chức năng thụ tinh. Đây là một bước quan trọng cuối cùng để hoạt hóa tinh trùng tuyệt đối phải có ion Ca2+ để phản ứng xảy ra. Những thay đổi của phản ứng cực đầu cho phép các ion xâm nhập vào màng bào tương, thay đổi nồng độ ion Ca2+ tự do nội bào và kiềm hóa bào tương.
Khi tinh trùng tiếp xúc với noãn, phản ứng cực đầu xảy ra. Đầu tiên, màng bên ngoài acrosome và màng bào tương hòa nhập với nhau ở một số điểm ở nửa trước đầu tinh trùng. Sự hòa nhập giữa hai màng này tạo nên những khoảng trống (cổng ra) lớn dần và các chất chứa bên trong acrosome có thể thoát ra ngoài. Các acrosome chứa ít nhất 12 enzyme khác nhau và nhanh chóng xẹp đi sau khi các enzyme trên thoát màng.
(a) Acrosome ở tinh trùng chưa bị kích thích, màng bào tương nguyên vẹn
(b) Phản ứng cực đầu, các hạt trong acrosome được phóng thích;
(c) Tinh trùng mất acrosome và chuẩn bị xâm nhập màng zona.
Trong số những enzyme được biết của acrosome thì quan trọng nhất là hyaluronidase và một enzyme giống trypsin giàu năng lượng là acrosin, ngoài ra còn có một số enzyme khác như phosphatase arylsulfatase, collagenase, phosphorylase, phospholipase C và B-galactosidase. Trong acrosome, men acrosin được dự trữ ở dạng pro-acrosin, chi khi có phản ứng cực đầu nó mới được giải phóng và chuyển dạng hoạt động - acrosin. Hai enzyme này đều đóng vai trò quan trọng đối với sự xâm nhập của tinh trùng vào noãn. Hyaluronidase giúp làm tan chất acid hyaluronic giữa các cumulus bao quanh noãn. Sự hoạt hóa các thụ thể tinh trùng do ZP3 trên màng zona dẫn đến sự phosphoryl hóa protein và tăng Ca” nội bào. Các đường truyền tín hiệu dẫn đến sự phosphoryl hóa protein và phản ứng cực đầu gồm: (1) đường truyền adenylate cyclase - cAMP - proteinkinase A; (2) đường truyền phospholipase C - diacylglycerol - proteinkinase C; và (3) đường truyền guanylate cyclase - cGMP - protein G.
1.2 Sự tương tác giữa tinh trùng và màng thấu quang của noãn
1.2.1 Gắn kết tinh trùng - màng thấu quang
Màng thấu quang được cấu tạo là 1 lớp áo glycoprotein dày 15 um do noãn bào tiết ra với 3 glycoprotein chính là ZP1, ZP2 và ZP3. Sự gắn kết giữa bề mặt màng thấu quang với thành phần đặc hiệu đã được hoạt hóa của màng bào tương của tinh trùng có thể là một sự liên kết phụ thuộc calcium và rất cần thiết cho sự xâm nhập của tinh trùng qua màng thấu quang. Sự gắn kết này cần có sự hiện diện của receptor trên màng bào tương tinh trùng dù có phản ứng cực đầu hay không, nhưng chưa thể khẳng định cấu trúc tinh trùng có cần thiết cho sự gắn kết này hay không.
1.2.2 Di chuyển qua màng thấu quang
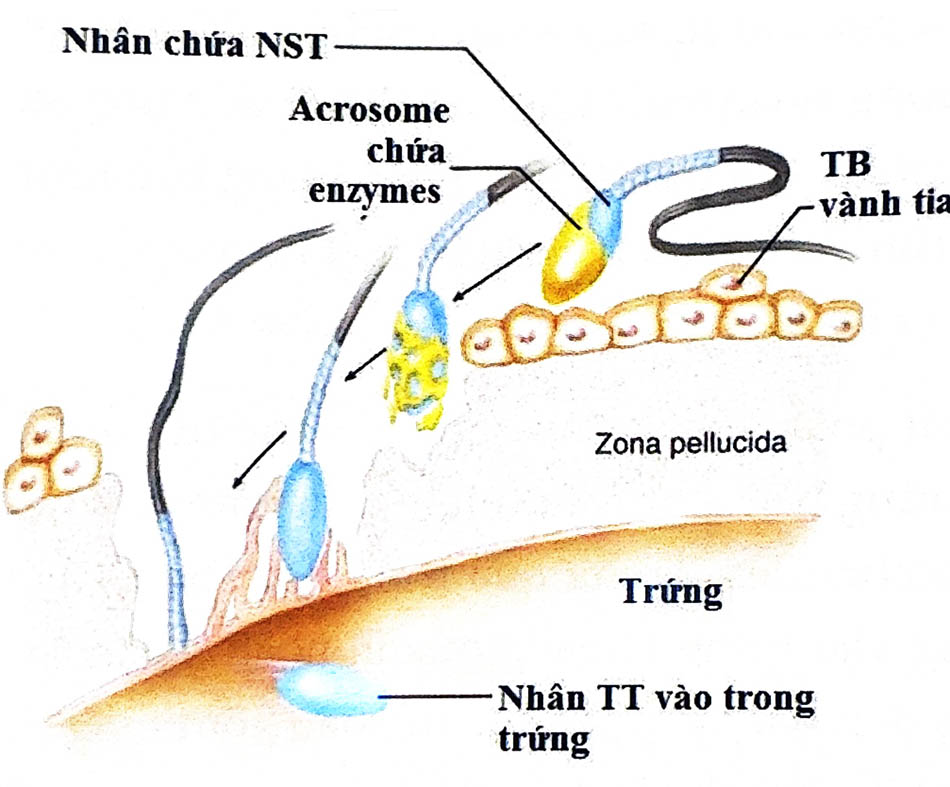
Bước gắn kết đặc hiệu như trên tạo cơ sở vững chắc cho tinh trùng chui vào màng thấu quang. Màng thế cực đầu đã phản ứng còn gắn lại ở bề mặt màng thất quang. Tinh trùng thường chui vào màng thấu quang theo một đường hẹp và cong dưới kính hiển vi điện tử, nó tạo với đường tiếp tuyến bề mặt noãn một góc khoảng 450 Nhưng đôi khi tinh trùng có thể xâm nhập theo hướng thẳng đứng hay nằm ngang s với bề mặt noãn. Cùng với lực chuyển động cần thiết của đuôi, acrosin đóng một vai trò quan trọng trong sự xâm nhập của tinh trùng. Thật sự acrosin được xem như là một “enzyme xâm nhập màng thấu quang” cho dù có thể cần phối hợp với những enzyme khác.
Tóm tắt các bước quan trọng trong sự xâm nhập của tinh trùng:
Màng bên ngoài của thể cực đầu hòa vào với màng bào tương tinh trùng.
Các hạt trong thể cực đầu vỡ ra giải phóng Lysine, các enzyme này “khoan” một đường qua màng ZP hoặc tạo sự thay đổi để các tiểu quản của acrosome có thể xâm nhập.
Màng thể cực đầu bên trong kéo dài tạo thành các tiểu quản do sự đa trùng hợp các actin hay các sợi giống actin nằm ở vùng sau thể cực đầu.
Khi đỉnh của các tiểu quản chạm đến màng bào tương noãn, hai màng này sẽ hòa nhập vào nhau.
1.3 Sự hòa nhập giao tử và hợp nhất tinh trùng vào noãn
Sau khi xuyên thủng và chui qua màng zona, tinh trùng nằm trong khoang quanh noãn và chờ một thời gian trước khi bám vào bề mặt màng bào tương noãn. Quan sát trên kính hiển vi điện tử thấy rõ ràng phần giữa đầu tinh trùng tương ứng với phần xích đạo hay vùng sau thể cực đầu tạo sự hòa nhập với noãn. Phần màng bào tương này che lên phần xích đạo của tình trùng khi tiếp xúc, tạo ra sự hòa nhập. Điều này cho thấy tầm quan trọng của vùng sau thể cực đầu được biệt hóa này. Tính ổn định đặc biệt của nó nhờ vào các liên kết protein cấu trúc giữa các màng song song, giúp nó không bị hòa tan trong phản ứng cực đầu. Xu hướng hòa màng này không biểu hiện nếu phản ứng cực đầu chưa xảy ra.
Quan sát trên kính hiển vi điện tử cũng cho thấy sự hòa nhập liên quan đến sų tương tác ban đầu với các vi nhung mao trên màng bào tương noãn. Sự hòa nhập này gây nên một số hiện tượng khác diễn ra trong vài phút: phản ứng vỏ, hoạt hóa noãn, hình thành và tống xuất cực cầu II. Ngay khi hòa nhập vào noãn, màng nhân tinh trùng biến mất, bảo tương noãn xuất hiện và chell bao lưng sau nhân của trùng giúp nhấn chìm phần đầu tinh trùng. Phần màng bào tương che phủ vùng đi phía sau và đuôi tinh trùng còn nằm lại trên bề mặt noãn.
1.4 Phản ứng vỏ
Noãn bào có sự tái phân bố các bào quan của bào tương để chuẩn bị cho hợp chuyển thành phôi. Những bào quan như những túi nằm trong phức hợp Golgi, chỉ enzym và mucopolysaccharides. Trong quá trình hoạt hóa, các hạt vỡ ra giải phóng các chất chứa bên trong vào trong khoang quanh noãn. Hiện tượng lan truyền ngoại bào của các hạt vỏ dẫn đến hai tác động tức thì, bao gồm tăng thể tích khoang quan noãn và màng zona chuyển thành cấu trúc dày cứng để bảo vệ. Phản ứng vỏ đám bà sự thụ tinh bình thường một tinh trùng thụ tinh với một noãn, tránh hiện tượng đa tinh trùng thụ tinh.
Hiện tượng lan truyền ngoại bào bắt đầu khoảng 10 giây sau khi tinh trùng xâm nhập vào zona. Thời gian trì hoãn này gọi là giai đoạn tiềm tàng. Các hạt tại vùng cạnh vị trí tinh trùng xâm nhập bị phá vỡ đầu tiên và tạo nên một làn sóng lan rộng chậm xung quanh bề mặt noãn bào. Các mucopolysaccharides được giải phóng từ cá hạt vào trong khoang quanh noãn gây nên sự tràn dịch nhanh chóng làm căng phồng màng thấu quang và giãn cách 1 – 20 km so với bề mặt noãn bào. Diễn tiến dạng sóng của sự lan truyền ngoại bào cùng với sự nâng lên của màng zona mất khoảng 20 gia và lan rộng quanh bề mặt noãn bào. Hơn 5 phút sau, khoang quanh noãn tiếp tục tăng lên và thấu quang dày hơn, cứng hơn.
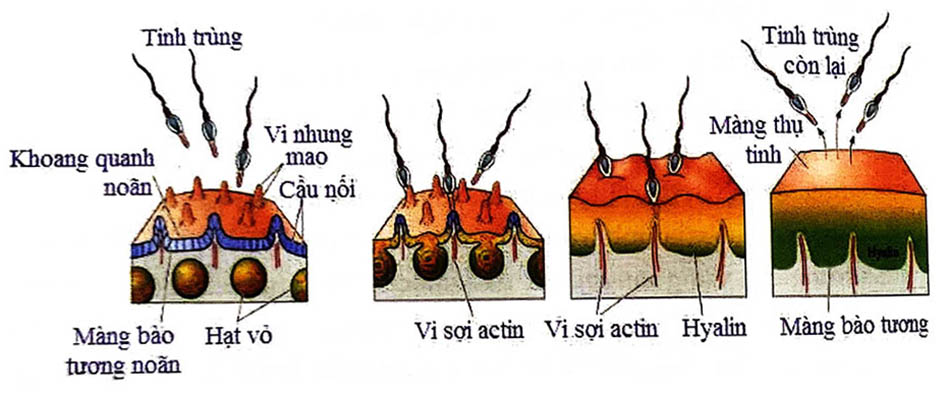
Yếu tố quan trọng của phản ứng vỏ là sự tăng Ca2+ trong bào tương. Ngay khi phát hiện thụ tinh, có sự giải phóng một lượng Ca2+ từ nơi lưu trữ nội bào, gây nên sự tăng Ca2+ từ 0.1 uM đến 10 pM. Trong vòng vài phút, Ca2+ trở về vị trí nghi. Vùng tăng Ca2+ bắt đầu tại điểm hòa nhập tinh trùng - noãn và truyền ngang qua noãn với những sóng tương tự nhỏ hơn, tiếp tục tồn tại nhiều phút sau khi thụ tinh. Cơ chế của sự lan truyền sóng chưa rõ. Những sóng Ca này tái diễn và kéo dài trong khoảng thời gian 3 phút; người ta có thể gây ra sóng này bằng cách tiêm các yếu tố tinh trùng hòa tan được vào trong noãn.
2 HOẠT HÓA NOÃN VÀ HỢP NHẤT TIỀN NHÂN
2.1 Hoạt hóa noãn
Sự di động của tinh trùng trong khoang quanh noãn nhanh chóng bị ức chế do hiện tượng hòa nhập tinh trùng. Lúc này, có sự thay đổi điện tích bề mặt noãn, phản ứng vỏ xảy ra ngăn hiện tượng đa thụ tinh. Sự hòa nhập kích thích hình thành thoi vô sắc giai đoạn trung kỳ giảm phân II của noãn thứ cấp, các NST kép tách nhau ra ở tâm động đi về hai cực tế bào. Một nửa bộ NST sẽ được tổng xuất ra ngoài noãn bào vào trong khoang quanh noãn theo thể cực thứ hai, còn lại noãn bào với bộ NST đơn bội.
Noãn trưởng thành (MII) chứa MPF (M-phase Promoting Factor - yếu tố thúc đẩy phase M) đã ổn định và hoạt hóa. Đây một yếu tố kiểm soát chu kỳ tế bào cần thiết, là một protein-serin/threonine kinase, gồm có cdc2 và cyclin B; và được ổn định bởi c-mos proto-oncogene là một yếu tố ổn định tế bào. Sự hoạt hóa và bất hoạt MPF xảy ra tùy thuộc vào sự phosphoryl hóa hay dephosphoryl hóa, phối hợp với các phân tử chu kỳ tế bào chính. Noãn thu được sau chọc hút trong IVF, tăng dần tỷ lệ MII sau khi cấy 4 - 6 tiếng và tình trạng hoạt hóa noãn duy trì cho đến khi tinh trùng xâm nhập. MPF bị bất hoạt khi đầu tinh trùng bị rã và hình thành màng nhân bao quanh tiền nhân đực.
2.2 Sự hình thành và hợp nhất tiền nhân
Khi đầu tinh trùng đi vào bào tương noãn, ngay lập tức màng nhân bị tan ra và chất nhân bị rã. Màng nhân tinh trùng bị tan ra do sự hòa lẫn lớp trong và lớp ngoài. Sự phồng lên của chromatin tinh trùng cô đặc cần thiết phá vỡ các liên kết chéo disulfide, nhờ tác dụng của yếu tố phát triển tiền nhân đực. Nhân tinh trùng lớn lên thành một mạng lưới các giải nucleoprotein, các protamine giàu Cystein có hàm lượng cao Arginine biến mất khỏi vị trí kết hợp với DNA tinh trùng. Sự trải rộng chromatin tinh trùng để hình thành tiền nhân có thể quan sát được nhờ vào sự hình thành màng nhân và sau đó là sự hiện diện của các hạt nhân con cô đặc. Sự hình thành tiền nhân đực phải được diễn ra trong điều kiện đặc hiệu của bào tương noãn giai đoạn trưởng thành, không thể xảy ra khi noãn chưa trưởng thành. Tiền nhân đực tích cực hơn trong sự hình thành tiền nhân. Sự sao chép DNA diễn ra trong thời gian từ 5 - 8 giờ sau khi tinh trùng xâm nhập.
Sự hình thành tiền nhân cái tương đối ít phức tạp hơn do chromatin của noãn không bị cô đặc cao như ở tỉnh trùng và không cần trao đổi protein nhân. Phần chromatin còn lại sau khi thể cực thứ hai bị tổng xuất được trải rộng và nó chịu sự thay đổi tương ứng để trở thành tiền nhân cái với màng nhân bọc ở ngoài, tương tự như ở tế bào sinh dưỡng bình thường. Tuy nhiên, đến giai đoạn cuối, siêu cấu trúc hai tiền nhân tương tự nhau và không còn có thể phân biệt tiền nhân đực và tiền nhân cái. Tiền nhân đực và tiền nhân cái được hình thành lúc này di chuyển đến gần nhau rồi cùng di chuyển đến trung tâm của noãn. NST của tinh trùng và noãn trộn lẫn vào nhau, tạo thành trung kỳ của lần nguyên phân đầu tiên.
3 SỰ PHÁT TRIỂN PHÔI CÁC GIAI ĐOẠN
3.1 Phôi giai đoạn phân cắt
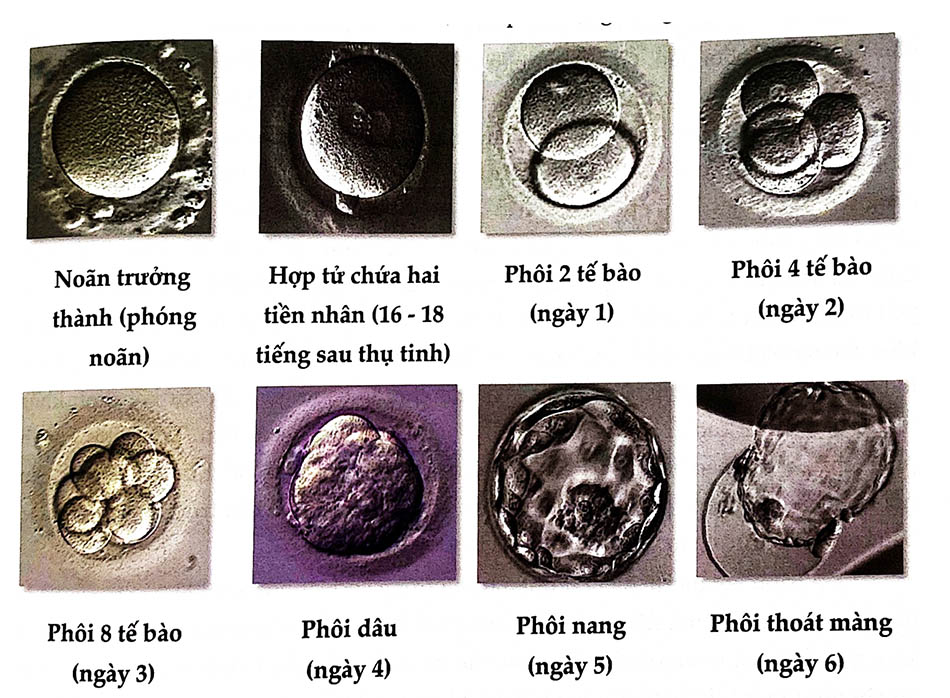
Sau khi hai tiền nhân tiếp cận và hòa nhập vào nhau, hợp tử ở vào giai đoạn trung kỳ của lần phân chia đầu tiên. Quan sát dưới kính hiển vi vào thời điểm 18 giờ sau thụ tinh để tìm hai tiền nhân (và thường kèm hai thể cực) là dấu hiệu chắc chắn thụ tinh đã xảy ra. Sự phân chia lần I kết thúc sau thụ tinh 24 giờ tạo thành phôi 2 tế bào, là chu kỳ kéo dài nhất, các chu kỳ sau chỉ khoảng 18 giờ. Những phân chia này kiểu như nguyên phân của tế bào bình thường các tế bào con được tạo ra gọi là các phôi bào. Phân chia lần II kết thúc sau thụ tinh 40 giờ, tạo thành phôi với 4 phôi bào kích thước tương đương nhau. Vào ngày 3, phôi chứa 6 - 12 phôi bào và ngày 4 gồm từ 16 - 32 tế bào.
Tuy nhiên, giai đoạn phân tách ngược lại với sự sinh noãn khi nó chỉ nhân đôi rồi phân chia NST và phân tách tế bào mà không có sự lớn lên (sự sinh noãn chỉ lớn lên chứ không nhân đôi và không phân chia). Đối với các tế bào sinh dưỡng, sau mỗi lần nguyên phân, nó phải tổng hợp bào tương để tăng kích thước trước khi có thể tiếp lần nguyên phân tiếp theo, để duy trì kích thước ổn định. Các phôi bào con được sinh ra kích thước chỉ bằng một nửa tế bào ban đầu và chúng vẫn tiếp tục phân chia nhỏ hơn và nhỏ hơn. Đây là điểm khác biệt rất quan trọng của giai đoạn phân tách.
Càng về sau, sự khác biệt giữa các phôi bào càng tăng lên do sự phân chia các thành phần bào tương không đồng đều hoặc do những thay đổi diễn ra trong bản thân phôi bào khi phát triển. Nhân của mỗi phôi bào sẽ được định hướng theo môi trường bào tương khác nhau và vì thế ảnh hưởng lên hoạt động genome cũng khác nhau. Kết quả là mỗi phôi bào sẽ thiết lập cho riêng nó một chương trình phát triển và sẽ tăng dòng tế bào đặc hiệu. Những lần phân chia đầu tiên thường đồng bộ, nhưng những lần sau thì không. Các phôi bào được tổ chức thành từng nhóm hay từng lớp và mỗi nhóm này có tốc độ phân tách đặc trưng riêng. Đây là những cơ sở cho sự hình thành và biệt hóa cơ quan sau này như thần kinh, cơ, xương...
Kích thước toàn bộ của phôi không thay đổi gì trong giai đoạn phân tách mà vẫn giữ nguyên hình dạng của màng zona. Nuôi dưỡng cho phôi giai đoạn này do tự bản thân các phức hợp tế bào hạt - vành tia bao quanh nó và dịch tiết của biểu mô vòi tử cung. Protein dịch tiết vòi tử cung biểu hiện dạng điện chuyển như trong huyết thanh, kết hợp với ß-glycoprotein.
3.2 Giai đoạn phôi dâu
Sau một vài lần phân chia, phôi chứa từ 16 - 32 phôi bào có hình dáng như trái dâu nên gọi là phôi dâu. Giai đoạn này phôi đã xuống đến buồng tử cung. Sau lần phân chia thứ ba, trong phôi diễn ra quá trình kết đặc tế bào. Hiện tượng kết đặc tế bào xảy ra vào khoảng ngày thứ 3 - 4 làm cho các tế bào áp sát vào nhau, tăng tối đa sự tiếp xúc giữa các tế bào, khoang gian bào giảm đi, các đường viền quanh tế bào cũng mò đi và ranh giới giữa các tế bào lúc này trở nên khó phân biệt. Giai đoạn này có gia tăng đáng kể tổng hợp protein và DNA. Quá trình kết đặc tế bào phụ thuộc vào Calci và liên quan đến khung tế bào và phân tử uvomorulin kết dính tế bào (còn gọi là yếu tố kết dính cadherin). Lúc này, các phôi bào trở nên phân cực hóa rất cao, tạo điều kiện cho sự biệt hóa tế bào, các phôi bào chia thành 2 nhóm: nằm ở trung tâm là khối tế bào trong (ICM) và một lớp bao quanh là khối tế bào ngoài. Do sự kết dính khác nhau, mặt ngoài các tế bào trở nên lồi và mặt trong trở thành lõm. Trong quá trình phân chia, các tế bào bên ngoài có được các đặc tính của biểu mô, hình thành các cầu nối gian bào, tạo nên ranh giới chặt chẽ rất khó tách rời. Nhờ cấu trúc này, lớp tế bào bên trong sẽ được ngăn tiếp xúc với môi trường bên ngoài.
Quá trình kết đặc tế bào rất quan trọng trong sự biệt hóa khối tế bào trong và tế bào lá nuôi, quyết định đến sự hình thành phôi thai. Nếu hiện tượng này bị ức chế do kháng thể kháng uvomorulin, thi phôi vẫn có thể phát triển thành phôi nang nhưng nó chỉ gồm một túi các tế bào lá nuôi mà không có khối tế bào trong và vì thế không thể phát triển thành phôi sống được. Các nguyên bào phôi giai đoạn này đều có hình tròn, thống nhất về mặt hình thái, sinh hóa, tiềm năng phát triển. Tất cả các nguyên bào phôi đều có thể tham gia vào sự hình thành tổ chức phôi, chúng có tính toàn năng. Tuy nhiên, về sau, các đặc điểm này của phôi bào thay đổi đi. Màng thấu quang trong giai đoạn phôi dâu vẫn nguyên vẹn tạo điều kiện cho sự kết đặc tế bào và ngăn hai phôi (nếu có) nhập vào nhau. Nếu lớp tế bào trong phân chia ở giai đoạn sớm này, có thể phát triển thành song thai cùng hợp tử.
3.3 Giai đoạn phôi nang
Phôi dâu nằm trong lòng tử cung được vùi trong niêm dịch của tử cung. Với sự hấp thụ của phôi, chất dịch này thấm qua màng thấu quang vào giữa các phôi bào tạo thành những túi dịch nhỏ. Các túi dịch tăng dần kích thước, lớn dần và sáp nhập vào nhau tạo nên một khoang lớn là khoang phôi nang. Do ranh giới giữa các tế bào bên ngoài chặt, dịch tiếp tục đi vào chủ yếu nằm giữa các tế bào của khối tế bào bên trong. Khối tế bào bên trong bị chèn tạo thành khối cô đặc và bị đẩy về một phía của khoang phôi. Khối tế bào bên ngoài trở thành một lớp biểu mô mỏng. Nó sẽ phát triển thành lá nuôi để tạo nên bánh nhau và các phần phụ của thai, còn khối tế bào trong chính là mầm phôi sẽ phát triển thành cơ thể phôi thai. Nơi mầm phôi nhô vào bên trong lòng khoang phôi nang gọi là cực phôi, đối diện với cực phôi là cực đối phôi. Các tế bào của phôi nang khác biệt nhiều so với các tế bào trong những lần phân chia đầu. Tuy nhiên, sự khác biệt giữa lá nuôi và mầm phôi lúc này chưa đáng kể.
4 SỰ PHÂN CHIA TẾ BÀO PHÔI
Không giống như các tế bào cơ thể khỏe mạnh bình thường, phôi bào không cần các phân tử tín hiệu đặc trưng để khởi động sự phân chia. Liên quan đến sự điều hòa phân chia tế bào, phôi tự điều hòa quá trình này nhờ vào đồng hồ nội bào. Các giai đoạn phân chia như sau:
Phase G1: diễn ra sự tổng hợp ARN (sao mã) và tổng hợp protein. Trong suốt giai đoạn này, tế bào không phân chia. Thời gian kéo dài khác nhau của giai đoạn này sẽ quyết định tốc độ phân chia tế bào. Sự tổng hợp DNA xảy ra ở phase S.
Phase G2: là phase chuyển tiếp từ phase S sang phase M. Phase M là giai đoạn nguyên phân, là lúc bắt đầu cô đặc và lúc kết thúc phân rã NST. Cô đặc là một quá trình cần thiết cho sự hình thành cấu trúc NST đặc hiệu của DNA. Sau khi hoàn tất phân rã NST, tế bào trở lại phase G1. Chu kỳ tế bào được điều hòa ở 3 điểm, không phụ thuộc vào sự phân chia nguyên phân hay giảm phân: (1) quá trình khởi động ở phase G1, (2) trước khi bắt đầu phase M, (3) giai đoạn trung kỳ. Phosphoprotein pp34 đóng một vai trò thiết yếu trong 3 quá trình tế bào này, kể cả tế bào cơ thể bình thường và tế bào phôi.
Trước khi bắt đầu nguyên phân, pp34 phóng thích nhóm phosphorus và kết hợp với cyclin, một protein chính yếu khác trong điều hòa chu kỳ tế bào. Toàn bộ phức hợp pp34-cyclin được gọi là yếu tố thúc đẩy trưởng thành MPF. Khi cyclin trong phức hợp này được phosphoryl hóa sẽ hoạt hóa enzyme và đảm bảo được sự cô đặc nhiễm sắc thể để bắt đầu phase M. Tác động kinase của phức hợp MPF cần thiết cho sự cô đặc nhiễm sắc thể. Như vậy, protein trong các nhiễm sắc thể chịu sự phosphoryl hóa của dưới ảnh hưởng của MPF. Tuy vậy, MPF không ổn định, nó hoạt hóa một protease làm phá vỡ đơn vị cyclin trong phức hợp MPF-cyclin, làm bất hoạt MPF. Sau đó, các tế bào đi vào trung kỳ nguyên phân và NST phân rã. Trong phase G1, một cyclin đặc hiệu với G1 kết hợp với pp34 tạo ra phase S. Sau khi phức hợp này phân ly, chu kỳ tế bào lại có thể bắt đầu.
Bình thường sự phân chia bào tương và phân chia nhân diễn ra đồng đều, NST sẽ được chia đều cho hai tế bào con. Khi sự phân chia này không đồng đều sẽ tạo ra các phôi bảo có bộ NST bất thường, phôi bào có nhiều nhân hoặc không có nhân. Các bào tương không nhân tiếp tục phân chia cho đến khi chỉ còn lại một mảnh bào tương không nhân.
5 BIỂU HIỆN GEN VÀ SỰ TỔNG HỢP PROTEIN CỦA PHÔI
Trước khi thụ tinh, noãn trưởng thành đã có cấu tạo sinh hóa hoàn chỉnh cho sự tổng hợp protein. Trong giai đoạn lớn lên, noãn đã chứa đủ các ribosome (bào quan tổng hợp protein), ARN ribosome (rARN), ARN thông tin (mARN) và ARN vận chuyển (tARN). Trong chu kỳ tế bào đầu tiên sau thụ tinh, protein được tổng hợp, điều chỉnh và bị phá vỡ vào các thời điểm cụ thể. Quá trình này vẫn tiếp tục trong thực nghiệm kể cả khi lấy nhân ra khỏi phối và khi có các tác nhân ức chế đặc hiệu sự tổng hợp ARN. Điều này cho thấy, trong những chu kỳ tế bào đầu tiên, sự chuyển hóa và tổng hợp protein hoàn toàn chịu sự kiểm soát của thông tin mà noãn đã nhận được từ mẹ trong quá trình phát triển và trưởng thành.
Các thông tin di truyền của phôi, kể cả phần xuất phát từ bố chỉ được dùng ở giai đoạn sau. Điều này xảy ra sau khi hoạt hóa genome của phôi, gọi là sự chuyển tiếp thông tin từ mẹ sang hợp tử (MZT). Mặc dù những kỹ thuật sinh hóa và sinh học phân tử nhằm phát hiện MZT chưa được thực hiện trên phôi người, nhưng dựa vào những thay đổi hình thái nhân chứng tỏ sự chuyển thông tin di truyền từ mẹ sang phôi diễn ra vào khoảng giai đoạn 8 tế bào.
Nghiên cứu dạng protein khi thông tin mẹ truyền sang phôi cho thấy, một nhóm protein có trọng lượng phân tử giống nhau từ 65.000 - 70.000 Dalton được tạo ra đầu tiên. Nhóm này gọi là protein yêu cầu sao mã (TRPs) và biến mất sau giai đoạn 8 tế bào. Khi sự tổng hợp DNA và phân chia tế bào trong phôi đã bị ức chế hoàn toàn thì lúc này TRP vẫn còn hiện diện. Điều này cho thấy sự giải mã được khởi đầu nhờ đồng hồ sinh học và chạy liên tục, không phụ thuộc vào sự tổng hợp DNA và phân chia tế bào.
Sự truyền thông tin từ mẹ sang phôi có ý nghĩa quan trọng về mặt sinh hóa và thực hành khi thực hiện nuôi cấy phôi. Trong thực hành, giai đoạn MZT diễn ra trùng với giai đoạn phôi ngừng phân chia nếu điều kiện môi trường thiếu sót. Hiện nay, người ta vẫn chưa hiểu điều kiện môi trường ảnh hưởng như thế nào đến MZT và liệu có liên quan đến các phân tử tín hiệu đặc trưng hay không.
Như vậy, sự phát triển của phôi trước khi làm tổ chỉ diễn ra trong khoảng 1 tuần đầu tiên sau thụ tinh. Hợp tử ban đầu trải qua giai đoạn phân chia làm tăng số lượng các phôi bào. Phôi dâu hình thành khoảng ngày 3 - 4 với đặc trưng hai khối tế bào bên trong và bên ngoài. Sự hình thành khoang phôi giữa các tế bào trong tạo nên phôi nang cùng với sự biệt hóa khối tế bào bên trong thành mầm phôi và biệt hóa khối tế bào bên ngoài thành lá nuôi. Chúng là cơ sở ban đầu cho sự hình thành cơ thể phôi cũng như bánh nhau và các phần phụ sau này. Phôi nang sẽ bắt đầu làm tổ ở nội mạc tử cung sau khi thoát khỏi màng thấu quang. Song song với những thay đổi về mặt hình thái là sự biến đổi mạnh mẽ về chuyển hóa, sinh tổng hợp protein và chuyển đổi gen. Vì thế, dù chỉ diễn ra trong thời gian ngắn, nhưng những thay đổi có tính quyết định của giai đoạn này làm cho nó đóng một vai trò vô cùng quan trọng trong sự sinh sản người.
6 SỰ LÀM TỔ CỦA PHÔI
6.1 Sự áp sát
Sự áp sát là hiện tượng tiếp xúc đầu tiên lỏng lẻo giữa phôi nang và nội mạc tử cung. Vị trí thường gặp trên nội mạc tử cung là các hốc nhỏ do khả năng tăng diện tích tiếp xúc với phôi nang đang có hình cầu. Đối với phôi nang, sau khi thoát màng thấu quang, có sự tiếp xúc trực tiếp giữa nguyên bào nuôi bên dưới và màng rụng của nội mạc tử cung. Gần như toàn bộ bề mặt của phôi nang có khả năng hình thành vị trí bám vào màng rụng.
Sự nhận diện các phân tử trên nguyên bào nuôi và biểu mô nội mạc tử cung làm trung gian cho sự tương tác ban đầu vẫn chưa hoàn toàn sáng tỏ. Tuy nhiên, protein được glycosyl hóa MUC1, một thành viên của họ Mucin, đóng vai trò quan trọng. MUC1 là một glycoprotein xuyên màng được biểu hiện ở đỉnh bề mặt của tế bào biểu mô nội mạc tử cung trong thời gian làm tổ ở người, làm trung gian cho sự gắn kết của phôi bào vào biểu mô tử cung.
6.2 Sự kết dính
Sự kết dính là sự gắn chặt của phôi vào nội mạc tử cung sau khi áp sát. Các nguyên bào nuôi kết dính bằng cách xâm nhập vào nội mạc tử cung nhờ vào các vị nhung mao. Nguyên bào nuôi có các sợi liên kết, Collagen type IV và các integrins hỗ trợ quá trình kết dính này.
MUC16 là một chất xuyên màng được hiện diện ở bề mặt đỉnh của biểu mô tả cung. Chất mucin này ngăn phôi bào làm tổ vào vị trí không mong muốn trên biểu mô. Như vậy, MUC16 ức chế sự kết dính tế bào.
Giao tiếp giữa phôi và nội mạc tử cung xảy ra ở giai đoạn này. Các tín hiệu truyền từ phôi nang đến nội mạc tử cung để tạo sự thích nghi tiếp theo. Chẳng hạn, những thay đổi trong bộ khung tế bào của các tế bào rụng giúp tách các tế bào khỏi liên kết với lớp đệm bên dưới, cho phép phôi bào xâm lấn sâu xuống dưới.
Thụ thể Proteoglycan: Một hệ thống thụ thể - phối tử khác tham gia vào quá trình kết dính là các thụ thể proteoglycan, được tìm thấy trên bề mặt màng rụng của tử cung. Các đối tác của chúng, các proteoglycan, được tìm thấy xung quanh các nguyên bào nuôi của phôi nang. Hệ thống thụ thể - phối tử này cũng hiện diện ở cửa sổ làm tổ.
6.3 Sự xâm nhập
Hợp bào nuôi (syncytiotrophoblast): Các phần nhô ra của các nguyên bào nuôi kết dính vào nội mạc tử cung tiếp tục tăng sinh và xâm nhập vào nội mạc tử cung. Khi các nguyên bào nuôi khi xâm nhập sẽ biệt hóa để thành một loại tế bào mới, hợp bào nuôi. Phần còn lại của nguyên bào nuôi gọi là tế bào nuôi (cytotrophoblast). Hợp bào nuôi không được xác định là một loại tế bào, đúng hơn là một mô đa nhân.
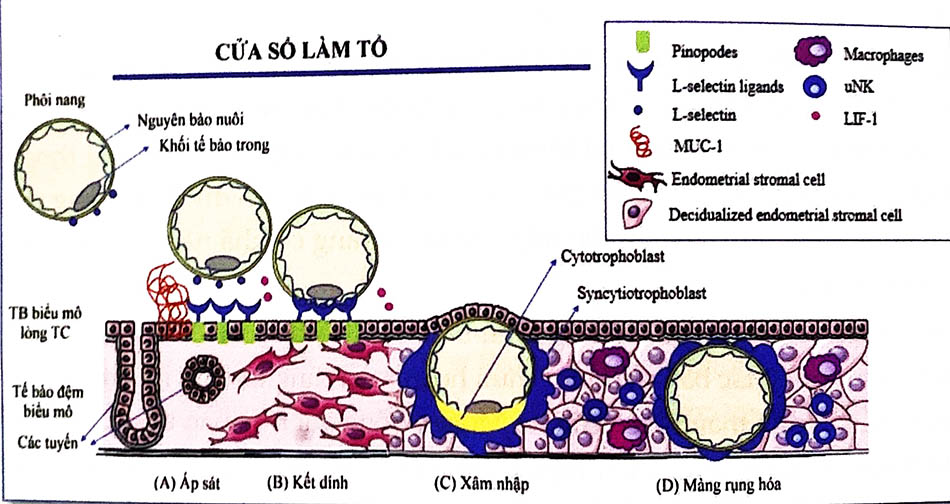
Sự xâm lấn tiếp tục với các hợp bào nuôi chạm đến màng đáy bên dưới các tế bào rụng, xuyên qua nó và tiếp tục xâm nhập vào lớp đệm tử cung. Cuối cùng toàn bộ phôi được chìm sâu vào nội mạc tử cung. Sau đó, các nguyên bào hợp bào tiếp xúc với máu mẹ và hình thành các nhung mao màng đệm. Đây là bước khởi đầu của việc hình thành nhau thai.
Sự xâm nhập của nguyên bào nuôi vào nội mạc tử cung được biểu hiện thông qua metalloproteinase MMP-2 và MMP-9. Hợp bào nuôi xâm nhập tử cung, tiếp cận nguồn cung cấp máu của mẹ, để thiết lập nền tảng cho tuần hoàn nuôi thai nhi.
Nguyên bào nuôi ngoài gai nhau (Extravillous trophoblasts): Là các tế bào từ nhung mao xâm nhập di chuyển vào cơ tử cung của mẹ. Những tế bào này tái cấu trúc các động mạch xoắn ốc để cải thiện và đảm bảo lưu lượng máu từ mẹ đến phôi thai đang phát triển. Quá trình này có thể xảy ra với các tĩnh mạch tử cung ổn định để cải thiện tuần hoàn thai nhi và các chất thải chuyển hóa. Các nguyên bào nuôi cũng đã được ghi nhận là có thể di chuyển vào cơ thể mẹ và đã được tìm thấy trong các mô khác nhau.
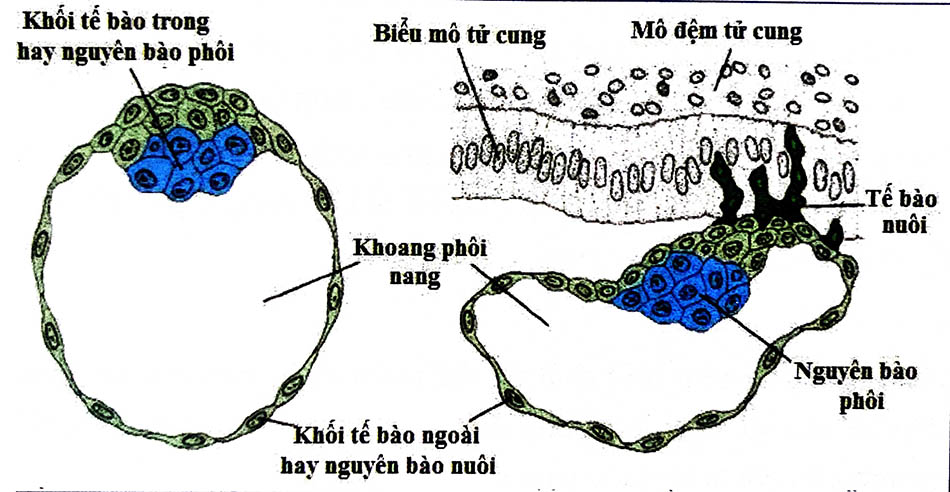
7 DIỄN BIẾN CHÍNH QUÁ TRÌNH LÀM TỔ
7.1 Sự chế tiết
Các phôi bào tiết ra các yếu tố đa mục đích trong quá trình xâm nhập. Một số yếu tố tự tiết, tự nhắm đích và kích thích chính nó xâm nhập sâu hơn vào nội mạc tử cung. Các chất tiết làm lỏng liên kết giữa các tế bào rụng ngăn cản sự đào thải phôi của cơ thể mẹ, thúc đẩy màng rụng hóa và ngăn hành kinh. Human Chorionic gonadotropin (hCG) là một yếu tố tăng trưởng tự tiết cho phôi nang. Yếu tố tăng trưởng giống Insulin 2 (IGF 2) kích thích sự xâm lấn.
7.2 Sự tách rời tế bào
Các hợp bào nuôi tách rời các tế bào rụng cả bằng cách thoái giáng các phân tử kết đinh tế bào liên kết các tế bào rụng với nhau cũng như thoái giáng chất nền ngoại bào. Các phân tử kết dính tế bào bị thoái giáng do hợp bào nuôi tiết ra yếu tố hoại tử u-alpha (TNF- a), gây ức chế sự biểu hiện của cadherins và beta-catenin. Cadherins là các phân tử kết dính tế bào và beta-catenin giúp gắn chúng vào màng tế bào. Do đó, liên kết giữa các tế bào rụng trở nên lỏng lẻo, cho phép các hợp bào nuôi và toàn bộ phôi xâm nhập vào nội mạc tử cung.
Chất nền ngoại bào bị phân hủy bởi serine endopeptidase và metalloproteinase. Các metalloproteinase như là collagenase, gelatinase và stromelysins gây tiêu hóa các type collagen và gelatin.
7.3 Ức chế miễn dịch
Phôi thai khác với các tế bào của người mẹ, và sẽ bị hệ thống miễn dịch của người mẹ đào thải nếu nó không tiết ra các chất ức chế miễn dịch. Một số tác nhân như yếu tố kích hoạt tiểu cầu (Platelet-activating factor), hCG, Prostaglandin E2, Interleukin 1-alpha, Interleukin 6, Interferon-alpha, yếu tố ức chế bạch cầu (leukemia inhibitory factor) và yếu tố kích thích khuẩn lạc (colony-stimulating factor).
Màng rụng hóa: các yếu tố từ phôi nang cũng kích hoạt sự hình thành cuối cùng của các tế bào rụng thành dạng phù hợp. Một số tế bào rụng ở gần phôi bào bị thoái hóa, cung cấp chất dinh dưỡng cho phôi.
7.4 Tắt kinh
HCG vừa hoạt động như một chất ức chế miễn dịch, vừa duy trì chức năng của hoàng thể, tiếp tục sản xuất các hormone sinh dục, giúp nội mạc tử cung tiếp tục phát triển, không hành kinh và là tín hiệu sớm nhất của thai kỳ.
8 TÀI LIỆU THAM KHẢO
- GS.TS. Nguyễn Vũ Quốc Huy,, GS.TS. Cao Ngọc Thành,, PGS.TS. Lê Minh Tâm,, PGS.TS. Trương Thành Vinh, cùng nhiều tác giả tham gia biên soạn, "Giáo trình Sản phụ khoa - Tập 1: Sản khoa", Đại học Huế - Trường ĐH Y Dược
- Cao Ngọc Thành, Lê Minh Tâm. Nội tiết phụ khoa và Y học sinh sản. Nhà xuất bản Đại học Huế, 2017.
- Aboubakr M. Elnashar, Gamal I. Aboul-Enein. Endometrial receptivity. Middle East Fertility Society Journal. 2004. 9(1): pp.10-24.
- Allais-Bonnet A, Pailhoux E. Role of the prion protein family in the gonads, Frontiers in Cell and Developmental Biology. 2014.
- Behre HM. Endokrinologie, Reproduktionsmedizin, Andrologie. Georg Thieme Verlag. 2002.
- Pansky B. Germ Layers and Their Derivatives. In: Review of medical embryology. 1982.
- Carlson B. Human Embryology and Developmental Biology. Sixth Edition. Elsevier. 2019.
- Clift D, Schuh M. Restarting life: fertilization and the transition from meiosis to mitosis, Nature Reviews Molecular Cell Biology. 2013. 14(9): pp.549-562.
- Linda ME, Eduardo D, Huff S. Color Atlas of Human Fetal and Neonatal Histology. Springer Nature Switzerland AG. 2019.
- Sadler TW. Langman's Medical Embryology. Thirteenth edition. Wolters Kluwer. 2019.
- Zhu, J.Y, Pang ZJ, Yu YH. Regulation of trophoblast invasion: the role of matrix metalloproteinases. Reviews in obstetrics & gynecology. 2012. 5(3-4): e137-e143.
