Sự phân chia tế bào sinh dục và sinh lý quá trình thụ tinh
1 Sự phân chia tế bào sinh dục
Cơ thể về cơ bản được bắt đầu phát triển bằng cách nhân lên và biệt hóa của các tế bào. Cơ thể người được tạo thành từ 60 - 100 nghìn tỷ tế bào, gồm hai loại: tế bào sinh dưỡng và tế bào mầm. Tế bào sinh dưỡng cần thiết cho sự tăng trưởng, phát triển, tái tạo và duy trì các mô khác nhau của cơ thể, trong khi tế bào mầm cần thiết cho việc sản xuất giao tử. Sự sống bắt đầu như một tế bào đơn lẻ - hợp tử - được hình thành bởi sự kết hợp của các giao tử đực và cái. Ở người, giao tử đực là tinh trùng được tạo ra ở tinh hoàn từ tuổi dậy thì, giao tử cái là tế bào noãn thứ cấp được phóng ra từ buồng trứng theo chu kỳ trong giai đoạn sinh sản
1.1 Đặc điểm nhiễm sắc thể
Mỗi tế bào cơ thể (tế bào sinh dưỡng) có bộ nhiễm sắc thể (NST) lưỡng bội, gồm 46 NST được sắp xếp thành 23 cặp, trong đó 22 cặp NST thường và cặp NST giới tính. Nhiễm sắc thể giới tính gồm hai loại: X và Y. Nữ giới có hai nhiễm sắc thể X trong khi nam giới có một nhiễm sắc thể X và một nhiễm sắc thể Y. Bộ NST người được biểu diễn bằng công thức 44,XX ở nữ và 44,XY ở nam.
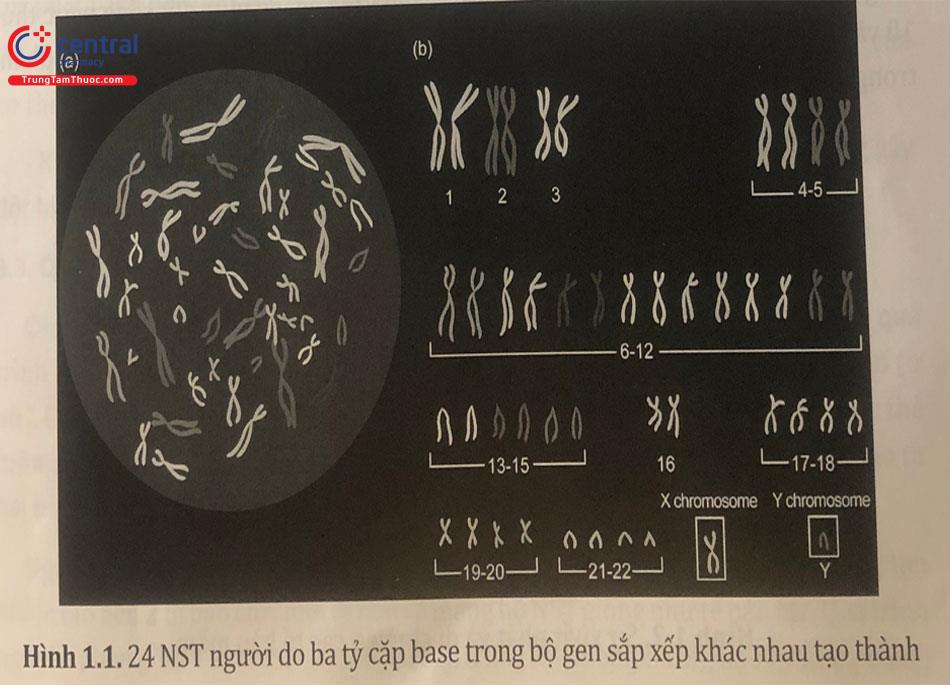
Giao tử là tế bào chuyên biệt để sinh sản. Mỗi tế bào giao tử có bộ NST đơn bội, gồm 23 NST. Ở nữ giới, tế bào noãn thứ cấp chỉ có một loại, gồm 22 NST thường và một NST giới tính X (22,X). Ở nam giới, tinh trùng có hai loại NST giới tính - NST X (22,X) và Y (22,Y).
Thành phần hoá học của NST chủ yếu là DNA và protein loại histone. Trong cấu trúc không gian, các phân tử ADN (chứa khoảng 140 cặp nuccleotide) quấn quanh các khối hình cầu protein (gồm 8 phân tử histon) tạo nên chuỗi nucleosome. Chuỗi nucleosome tạo thành sợi cơ bản đường kính khoảng 100Å. Sợi cơ bản xoắn bậc hai tạo thành sợi nhiễm sắc đường kính khoảng 250Å. Các sợi nhiễm sắc tiếp tục xoắn và bện lại theo nhiều cách tạo thành cấu trúc chromatid.
Các NST có thể phân loại theo chiều dài toàn bộ, vị trí tâm động, chiều dài mỗi cánh kể từ tâm động. Bình thường khi tế bào không phân chia, các NST rất dài, nằm trải rộng ra và chồng chéo nhau, không thể thấy dưới kính hiển vi quang học. NST có nhiều hình dạng khác nhau như hình hạt, hình sợi, hình que, hình V, dài từ 0,2 đến 50 um, đường kính 0,2 - 2 um.
1.2 Sự xuất phát và di chuyển của các tế bào mầm
Các tế bào mầm nguyên thuỷ là tiền thân sớm nhất của giao tử. Ở người, các tế bào mầm nguyên thuỷ xuất hiện ở ngoại phôi bì từ tuần thứ hai phát triển. Sau đó chúng tách ra và di chuyển vào túi noãn hoàng. Ban đầu, chúng nằm như một khối riêng ở trung bì ở đoạn cuối của phôi và sau đó là trong nội bì túi noãn hoàng vào tuần lễ thứ tư.
Từ tuần lễ thứ tư đến thứ sáu phôi thai, các tế bào mầm nguyên thuỷ tiếp tục di chuyển kiểu amíp từ túi noãn hoàng đến thành ruột, đi qua mạc treo ruột đến thành sau của lưng. Tại thành sau, các tế bào này nằm hai bên đường giữa, trong trung mô lỏng lẻo. Hầu hết các tế bào mầm gắn vào thành lưng ngang mức đốt sống ngực thứ 10 và sẽ trở thành tuyến sinh dục. Các tế bào mầm nguyên thuỷ liên tục nguyên phân trong suốt quá trình di chuyển.
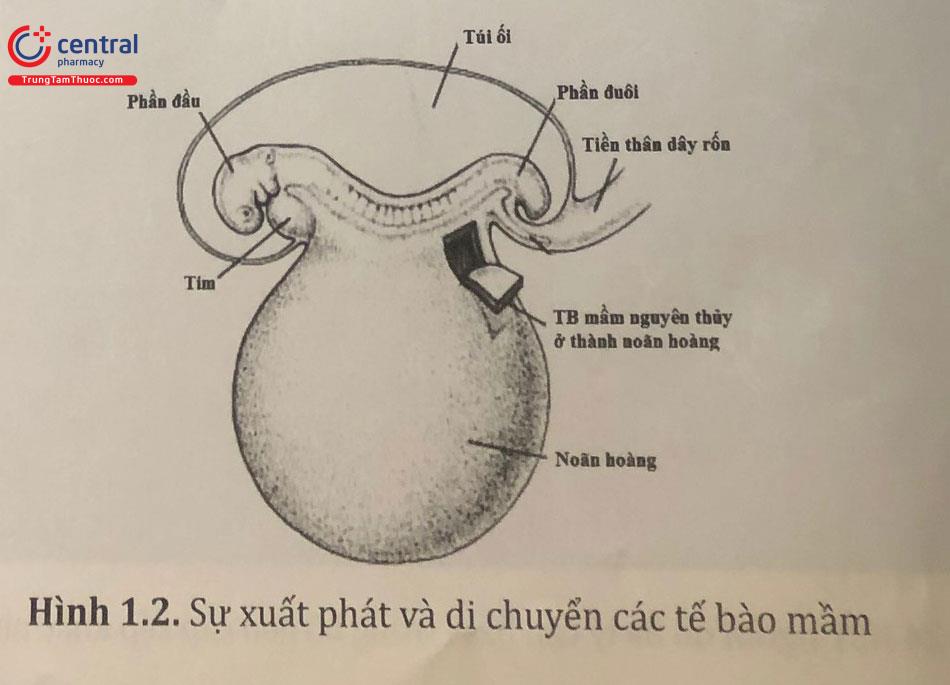
Khi các tế bào mầm di chuyển đến vùng sinh dục, chúng kích thích các tế bào biểu mô và trung thận - thận giai đoạn phôi tăng sinh và tạo thành dải mô gọi là thừng sinh dục nguyên phát. Thừng sinh dục tăng sinh thành dải sinh dục là biểu hiện của tuyến sinh dục nguyên phát. Thừng sinh dục tăng sinh các tế bào mầm, nuôi dưỡng và điều hoà sự phát triển các tế bào sinh dục - nang noãn ở nữ và tế bào Sertoli ở nam trong biểu mô mầm ở tiểu quản sinh tinh. Thừng sinh dục rất cần thiết cho sự phát triển của tế bào mầm. Nếu chúng không đến được vùng sinh dục, thì sự phát triển tuyến sinh dục không thể xảy ra.
1.3 Nguyên phân
1.3.1 Thời điểm nguyên phân
Ở nam giới, các tế bào mầm nguyên thuỷ sau khi di chuyển đến tuyến sinh dục sẽ ngủ yên cho đến tuổi dậy thì. Khi dậy thì, hoạt động nguyên phân tại biểu mô ống sinh tinh sẽ diễn ra liên tục suốt cuộc đời.
Khác với nam giới, ở nữ, noãn nguyên bào qua một giai đoạn hoạt động nguyên phân mạnh mẽ trong buồng trứng phôi thai từ tháng thứ hai đến tháng thứ năm của thai kỳ. Trong suốt giai đoạn này, mật độ tế bào mầm tăng lên theo cấp số mũ từ chỉ vài ngàn lên đến gần bảy triệu (khoảng 2 triệu noãn nguyên bào, 4,8 triệu noãn bào sơ cấp). Sau đó, số lượng các tế bào mầm giảm đi nhanh chóng còn khoảng 2 triệu vào lúc sinh, chủ yếu do quá trình thoái hoá. Vì vậy, số lượng noãn bào của bé gái lúc sinh ra là giới hạn, và chỉ giảm đi theo tuổi kể từ lúc dậy thì.
1.3.2 Ý nghĩa của nguyên phân
Nguyên phân tạo ra số lượng tế bào nhiều hơn từ nguồn tế bào ban đầu, giúp cơ thể phát triển hay thay thế cho các tế bào mất đi do bị chết hay tổn thương.
Duy trì số lượng NST lưỡng bội qua các lần phân chia, giúp ổn định bộ NST của cơ thể.
Không có sự trao đổi chéo hay tái sắp xếp các thông tin di truyền nên không gây đột biến.
1.3.3 Quá trình nguyên phân
Các tế bào mầm nguyên thuỷ tiếp tục nhân lên qua sự nguyên phân trong quá trình di chuyển. Những tế bào mầm nguyên phân tạo thành các noãn nguyên bào (ở nữ). Giống như những tế bào khác của cơ thể, noãn nguyên bào có bộ nhiễm sắc thể lưỡng bội, chứa 23 cặp NST. Trong một lần nguyên phân, mỗi noãn nguyên bào tạo ra hai bộ NST lưỡng bội giống hệt nhau về mặt di truyền.
Nguyên phân là hoạt động phân chia tế bào trải qua một loạt các khẩu phức tạp nhằm tạo nên 2 tế bào con, mỗi tế bào sẽ mang bộ NST giống như tế bào mẹ. Quá trình nguyên phân gồm 4 kỳ: tiền kỳ (prophase), trung kỳ (metaphase), hậu kỳ (anaphase) và chung kỳ (telophase). Thời kỳ tế bào không hoạt động phân chia gọi là gian kỳ (interphase).
Khi tế bào bắt đầu phân chia, NST kết đặc lại và nhân đôi. Kết quả là lúc bắt đầu giảm phân, các tế bào có 2n4c (trong cách biểu diễn này, n là số NST loài, và c là số chromatid trong một NST đơn trước khi diễn ra sao chép DNA). Vì vậy, mỗi NST lúc này chứa hai nhiễm sắc tử dính nhau ở tâm động. NST có dạng dài và mảnh. Màng nhân bị phá vỡ và hình ảnh của nhân không xác định được. Cũng trong tiền kỳ, các trung thể tách ra, mỗi trung thế tiến về mỗi cực tế bào.
Ở trung kỳ, các NST tiếp tục kết đặc lại và bắt màu thuốc nhuộm đặc hơn, tập trung trên mặt phẳng xích đạo của thoi vô sắc. Ở thời điểm này, có thể quan sát được NST rõ ràng nhất.
Ở hậu kỳ, hai nhiễm sắc tử của mỗi NST tách nhau ra ở tâm động và di chuyển về hai cực của tế bào. Lúc này mỗi nhiễm sắc tử gọi là NST con.
Khi NST đến mỗi cực của tế bào thì chung kỳ bắt đầu, các NST tháo xoắn, bắt màu thuốc nhuộm nhạt dần và màng nhân hình thành. Sự phân chia của bào tương bắt đầu với sự xuất hiện vòng thắt trên màng tế bào. Cuối cùng tế bào mẹ ban đầu tách ra thành hai tế bào con với bộ NST hoàn toàn giống nhau.
.jpg)
1.4 Giảm phân
1.4.1 Thời điểm giảm phân
Thời điểm giảm phân ở hai giới cũng khác nhau. Ở nam giới, các tế bào mầm nguyên thuỷ vẫn ngủ yên từ lúc 6 tuần tuổi thai kỳ cho đến khi dậy thì. Lúc dậy thì, các tiểu quản sinh tinh trưởng thành và tế bào mầm trải qua nguyên phân liên tục suốt cuộc đời song song với sự biệt hoá để thành tinh trùng thông qua hoạt động giảm phân.
Ở nữ giới, sau khi các tế bào mầm nguyên thuỷ trải qua một vài lần nguyên phân từ tháng thứ ba của thai kỳ, hoạt động giảm phân xảy ra song song với hoạt động nguyên phân. Có điều, khi vào tiền kỳ của giảm phân I (giai đoạn diplotene) các noãn bào sẽ bị ngưng trệ. Tình trạng này xảy ra với toàn bộ noãn bào ở buồng trứng cho đến khi dậy thì. Khi dậy thì, với sự phát triển của nang noãn dưới tác động nội tiết sinh sản, noãn bào sẽ vượt qua block và tiếp tục giảm phân đến giai đoạn trung kỳ của lần giảm phân II. Block lần thứ hai này chỉ được giải phóng khi noãn bào được thụ tinh với tinh trùng.
1.4.2 Tầm quan trọng về mặt sinh học của sự giảm phân
Giảm số lượng NST từ lưỡng bội (2n) thành đơn bội (n), như thế, số NST của loài có thể được duy trì từ thế hệ này qua thế hệ khác.
Sự tái sắp xếp độc lập của NST bố và mẹ (các NST tương đồng) tạo sự kết hợp các đặc tính di truyền tốt hơn.
Tái phân bố thông tin di truyền của bố và mẹ qua quá trình trao đổi chéo trong lần giảm phân thứ nhất.
1.4.3 Quá trình giảm phân
Tương tự như nguyên phân, mỗi lần giảm phân cũng gồm các giai đoạn khác nhau diễn tiến từ tiền kỳ đến trung kỳ, hậu kỳ, rồi đến chung kỳ. Khoảng thời gian trước giảm phân được gọi là Interphase (gian kỳ - pha chuẩn bị), đó là khi diễn ra sự sao chép DNA.
Khác với nguyên phân, sự giảm phân gồm hai lần phân chia, giảm phân I và giảm phân II. Trong lần giảm phân I các NST tương đồng tách ra, trong lần phân bào II các chromatid tách ra. Vì thế, lần giảm phân thứ nhất là sự phân chia giảm nhiễm, có tiền kỳ kéo dài, cho phép sự trao đổi thông tin di truyền giữa các cặp NST. Tiền kỳ được chia theo sự biểu hiện của nhân, dưới các tiểu giai đoạn:
- Leptotene (sợi mảnh): Đây là giai đoạn đầu tiên. Các NST cô đặc trở nên dài, có cấu trúc cuộn lại lỏng lẻo.
- Zygotene (tiếp hợp): Đây là thời điểm các đoạn tương đồng của NST bố và mẹ kết hợp tạo cặp. Sự kết cặp của các NST tương đồng được gọi là tiếp hợp (synapsis), và quá trình này dẫn đến một nhân chỉ có đơn bộ NST mặc dù tế bào vẫn còn 2n, 4c.
- Pachytene (sợi dày): Trong giai đoạn này, các NST dày lên và ngắn lại, và bắt đầu xảy ra sự trao đổi chéo, dẫn đến sự trao đổi các phần giữa các NST đã kết cặp. Mỗi cặp tiếp hợp được gọi là bivalent (thể lưỡng giá), người ta có thể xác định mỗi cặp lưỡng giá gồm 4 sợi nhiễm sắc (chromatids), gọi là tetrad (bộ bốn). ha tu GE GT C
- Diplotene (tách đôi): Các chromatids rõ hơn. Các chromatids con vẫn còn kết cặp chặt chẽ và nối kết với nhau ở tâm động nhưng các chromatids bố và mẹ đã bắt đầu đẩy xa nhau và tách ra. Tuy nhiên, chúng vẫn còn gắn với một điểm nào đó gọi là sự giao thoa (chiasmata). Mỗi sự giao thoa tiêu biểu cho một vùng nơi đó có sự trao đổi chéo (crossing-over), giữa các sợi nhiễm sắc nguồn gốc từ bố và mẹ.
- Diakinesis (hướng cực): Các NST rút ngắn và dày tối đa. Sự giao thoa bước vào giai đoạn cuối. Sự hướng cực đồng hành với sự biến mất của hạch nhân và mảng nhân (phá vỡ túi mầm), tạo nên thể hình thoi.
.jpg)
Trong trung kỳ của lần giảm phân I (metaphase I), các thể lưỡng giá xếp thẳng hàng trên mặt phẳng xích đạo của thoi vô sắc, nhờ đó vào thời điểm hậu kỳ, các NST tương đồng di chuyển về phía cực đối diện của thoi. Khác với nguyên phân, các tế bào con sau lần giảm phân thứ nhất nói chung là không giống nhau. Mỗi tế bào con của lần giảm phân thứ nhất chứa số NST đơn bội, nhưng mỗi NST vẫn còn chứa 2 nhiễm sắc tử nối với nhau ở tâm động.
Giữa lần giảm phân I và II không có sự sao chép DNA mới. Do thiếu gian kỳ, nên không có tiền kỳ nổi bật. Sự giảm phân lần II tương tự một sự phân chia bình thường. ngoại trừ tế bào mang NST đơn bội kép (1n, 2c). Khi các NST xếp hàng dọc theo mặt phẳng xích đạo lúc trung kỳ II, các tâm động giữa các nhiễm sắc tử chị em tách nhau ra ở tâm động cho phép các sợi nhiễm sắc của mỗi NST di chuyển đến các cực đối diện của thoi vô sắc trong hậu kỳ II. Mỗi tế bào con của giảm phân lần hai thật sự là tế bào đơn bội (1n, 1c).
1.5 Kết luận
Sự phân chia các tế bào trong cơ thể là một hoạt động hết sức tinh vi và phức tạp, trải qua thời gian dài trong cuộc đời con người và theo những quy luật nghiêm ngặt. Giảm phân là một đặc điểm phân chia quan trọng của giao tử, giúp đảm bảo khả năng sinh sản hữu tính thông qua việc giảm đi một nửa NST để mỗi tế bào mầm có số lượng nhiễm sắc thể đơn bội. Khi hai tế bào mầm hợp nhất để tạo thành hợp tử, số lượng nhiễm sắc thể được khôi phục lại bình thường. Giảm phân còn là cơ sở cho các biến dị di truyền. Do sự phân loại ngẫu nhiên của các nhiễm sắc thể của bố và mẹ, và sự trao đổi vật chất di truyền trong quá trình giảm phân nên các tế bào con (tức là giao tử) có thể có cấu hình di truyền mới. Điều này tạo ra các biến thể cá nhân trong loài, là điều cần thiết cho quá trình tiến hóa. Đồng thời, giảm phân tạo sức sống lai, giúp duy trì sức sống ở thế hệ con cái thông qua sinh sản hữu tính.
1.6 Tài liệu tham khảo
1. Bruce M. Carlson (2019). Human Embryology and Developmental Biology. Sixth Edition. Elsevier.
2. Cao Ngọc Thành, Lê Minh Tâm (2017). Nội tiết phụ khoa và Y học sinh sản. Nhà xuất bản Đại học Huế.
3. Linda M. Ernst, Eduardo D. Ruchelli Chrystalle Katte Carreon Dale S. Huff (2019). Atlas màu về mô học của thai nhi và trẻ sơ sinh. Springer Nature Thụy Sĩ AG
2 Sự sinh hoãn
2.1 Giới thiệu
Sự sản xuất giao tử ở nữ giới là một trong những quá trình phức tạp và kéo dài nhất. Quá trình sinh noãn bắt đầu từ thời kỳ bào thai, tiếp tục sau khi sinh, tăng lên vào giai đoạn dậy thì và đạt cực điểm vào thời điểm phóng noãn. Sự thay đổi quan trọng nhất là sự giảm phân, làm chia đôi số lượng nhiễm sắc thể (NST) trong khi sự thụ tinh làm khôi phục số lượng NST. Sự phóng noãn, phóng thích noãn từ buồng trứng vào đúng thời điểm thụ tinh và tiếp tục phát triển. Sự phóng noãn đòi hỏi phá vỡ nang De Graaf, dưới sự kiểm soát của hormon hướng sinh dục từ tuyến yên và tuỳ thuộc vào sự cân bằng nội tiết đặc biệt cần thiết cho sự lớn lên của nang noãn một cách đầy đủ. Giai đoạn cuối của giảm phân cần sự phát triển nang noãn hoàn chỉnh và phóng thích hormon của tuyến yên. Sự tương quan về thời gian của sinh noãn và phóng noãn thay đổi ở các loài khác nhau.
Sự sinh noãn là một quá trình rất quan trọng, đặc biệt trong đảm bảo tính ổn định và tiến hoá của loài qua sự di truyền. Quá trình này gồm sự hình thành, lớn lên và trưởng thành của giao tử cái, và có thể chia làm 4 giai đoạn:
- Các tế bào mầm nguyên thuỷ xuất phát từ bên ngoài phôi và di chuyển đến tuyến sinh dục, sau đó tăng số lượng qua nguyên phân.
- Giảm chất liệu di truyền - NST thông qua giảm phân.
- Trưởng thành về cấu trúc và chức năng của noãn.
- Sự hình thành và phát triển nang noãn: đảm bảo cho sự phát triển và biệt hoá của noãn, tạo môi trường ổn định xung quanh noãn cũng như hoạt động chế tiết hormone sinh dục giúp noãn trưởng thành, phóng noãn và giai đoạn hoàng thể.
.jpg)
Sự sinh noãn
Sự giảm phân của noãn Sự sinh noãn là quá trình hình thành giao tử cái - noãn bào từ các tế bào mầm nguyên thuỷ. Quá trình sinh noãn bắt đầu từ rất sớm, ở giai đoạn bào thai ở vỏ buồng trứng. Các tế bào mầm nguyên thuỷ phân chia nguyên phân để tạo thành một số lượng lớn các noãn nguyên bào. Mỗi noãn nguyên bào sau đó sẽ to ra để tạo thành một noãn bào sơ cấp. Noãn bào sơ cấp đi vào tiền kỳ phân chia giảm phân đầu tiên từ giữa giai đoạn bào thai. Nhưng sự phân chia này bị ngưng trệ cho đến tuổi dậy thì do sự hiện diện của yếu tố ức chế trưởng thành noãn bào (Oocyte maturation inhibitor - OMI) được tiết ra bởi các tế bào hạt bao quanh noãn. Lần giảm phân đầu tiên chỉ hoàn thành khi các noãn bào sơ cấp bắt đầu trưởng thành và chuẩn bị cho quá trình phóng noãn.
Ở tuổi dậy thì trong mỗi chu kỳ buồng trứng, 5 - 50 noãn bào sơ cấp được chiêu mộ và đi vào giảm phân khi có đỉnh LH vào giữa chu kỳ, ngay trước khi rụng trứng, tạo thành hai tế bào con với số lượng nhiễm sắc thể đơn bội. Giảm phân lần một là không cân đối; hầu hết tế bào chất đi đến một tế bào con tạo thành noãn bào thứ cấp, trong khi tế bào con kia nhận tế bào chất tối thiểu và hình thành thể cực thứ nhất.
Noãn bào thứ cấp đi vào giảm phân lần hai, nhưng lại ngưng trệ ở trung kỳ và chỉ kết thúc giảm phân khi có tinh trùng xâm nhập. Lần giảm phân hai cũng không cận bằng nên một tế bào con nhận được hầu hết tế bào chất, còn tế bào thứ hai nhận một lượng rất nhỏ tế bào chất và tạo thành thể cực thứ hai. Như vậy, một noãn bào nguyên phân chỉ tạo thành một noãn với 22 NST thường và một NST X; và ba thể cực.
.jpg)
2.1.1 Sự thay đổi ở tế bào chất
Trong giai đoạn phát triển của sự sinh noãn, ở tế bào chất diễn ra hoạt động tổng hợp và chuyển hoá rất mạnh, ảnh hưởng rất lớn đến kích thước của noãn. Đường kính noãn bào giai đoạn trưởng thành vào khoảng 120 um, lớn hơn nhiều so với tế bào sinh dưỡng (chỉ khoảng 10 um). Kích thước noãn bào phát triển đầy đủ chủ yếu do khối lượng dinh dưỡng trong bào tương, mặc dù kích thước nhân cũng phát triển. Kích thước này được duy trì cho đến khi trưởng thành hoàn toàn và ít có sự thay đổi đáng kể khác.
Hoạt động chủ yếu trong giai đoạn phát triển của noãn là sự tổng hợp RNA và protein, là các vật liệu di truyền hỗ trợ cho sự phát triển của phôi trong giai đoạn sớm sau thụ tinh. Các thành phần trong bào tương của noãn bào điển hình gồm yolk granules, pigment granules, và ty lạp thể. Hạt vỏ nằm ngay bên dưới màng bào tương. Hầu hết các noãn bào được bao quanh bởi một số lớp áo tế bào. Lớp trong cùng là lớp glycoprotein xơ đóng vai trò quan trọng trong sự tương tác giữa tinh trùng và noãn bào, được gọi là màng trong suốt.
Túi mầm trong noãn bào lớn lên, hạt nhân trở nên nổi bật và cấu trúc nhân của nó được xác định do sự tổng hợp ribosomal RNA bắt đầu. Ty lạp thể nhân lên và kéo dài với các nếp gấp ở màng trong ty lạp thể chạy ngang chuyển thành hình tròn hay hình bầu dục với các không bào. Chúng kết hợp chặt chẽ với lưới nội bào.
Phức hợp Golgi gồm các không bào, các túi lipid và khối các lá hình cung dẹt kết hợp với các túi và hạt được bao bọc bởi lipid. Nó tạo ra các hạt vỏ, sau này liên quan đến cơ chế bảo vệ, ngăn sự xâm nhập đa tinh trùng.
Noãn bào tổng hợp xấp xỉ 400 protein khác nhau, bao gồm a và B tubulin, B và actin, lactate dehydrogenase, ribosomal protein và histone, calmodulin và các loại khác. Một số trong các protein này được sản xuất ổn định qua các giai đoạn phát triển của noãn, một số khác có thể tích lũy từ huyết tương. Noãn bào tổng hợp thụ thể acetyl cholin màng và các tín hiệu đáp ứng gây ra do Ach điều hoà chuyển hoá phosphoinositide và chuyển động hoá ion calci nội bào. Những đáp ứng này có thể điều hoà sự tương tác tinh trùng - noãn và đa tinh trùng qua cơ chế cholinergic. Protein của màng trong suốt cũng được tổng hợp trong giai đoạn phát triển của noãn.
Ty lạp thể kết hợp với túi mầm và thoi vô sắc phân tách ở giai đoạn hậu kỳ. Nhiều vi quản có thể tạo thành “tiểu thể sao”. Các vị quản là các nhị trùng của tiểu đơn vị a và B và được cấu thành bởi các trụ rỗng đường kính 14 nm. Các vi quản hỗ trợ trong việc chuyển động của túi mầm trước khi sự trưởng thành bắt đầu, thiết lập ngăn chặn đa tinh trùng thụ tinh ở bào tương noãn bằng cách hướng cho các hạt vỏ di chuyển ra ngoại biên và làm tiền nhân có thể di chuyển trong noãn bào sau thụ tinh.
2.1.2 Trưởng thành về cấu trúc và chức năng của noãn
2.1.2.1 Sự trưởng thành tiền phóng noãn
Lúc bắt đầu sự trưởng thành tiền phóng noãn trong nang De Graaf, noãn vẫn còn là noãn bào sơ cấp. Khoảng ngày 13 - 14 chu kỳ kinh, thời điểm đạt đỉnh LH, noãn bào chứa các NST dạng chổi lông nhưng lúc này sự tổng hợp RNA chấm dứt. Các NST sau đó ngắn lại và dày lên, các vòng chổi của nó bị rút lại. Sự hướng cực xảy ra và hoàn thành giai đoạn tiền kỳ của giảm phân I. Quá trình giảm phân này tiếp tục phá vỡ túi mầm (15 giờ sau đỉnh LH) và dẫn đến sự di chuyển của các thể lưỡng giá đến vị trí dưới màng noãn. Trung kỳ I xảy ra sau đó và sự giảm phân I hoàn tất nhanh chóng, tạo hai tế bào con không giống nhau. Một tế bào lớn nhận hầu hết các bào tương, được gọi là noãn bào thứ cấp. Tế bào kia là một tế bào nhỏ không có chức năng gọi là thể cực thứ nhất. Ở người, thể cực này nhanh chóng thoái hoá trước khi phân chia lần một.
Ngay sau khi thể cực thứ nhất bị đẩy ra, noãn bào thứ cấp đi vào giảm phân lần II, nhưng lại một lần nữa quá trình giảm phân lại ngừng trệ, đây là thời điểm trung kỳ II (35 giờ sau đỉnh LH). Nói chung, sự phóng noãn xảy ra khi noãn bào thứ cấp ở giai đoạn trung kỳ II. Block lần thứ hai của giảm phân được giải thoát khi noãn được thụ tinh với một tinh trùng hay nói cách khác noãn bào thứ cấp không được thụ tinh sẽ không thể hoàn tất sự giảm phân lần II. Sau khi phóng noãn, phần nang còn lại tạo thành hoàng thể, và chính thể này sản xuất hormone steroids (lượng lớn progesterone). Sự phân chia tế bào chất của các tế bào con trong giảm phân lần II cũng không giống nhau; tạo ra noãn bào trưởng thành lớn và một thể cực thứ hai nhỏ không có chức năng.
2.1.2.2 Thay đổi về hình thái học
Sự trưởng thành diễn ra vào giai đoạn cuối của sự sinh noãn, với một số thay đổi diễn ra trong noãn bào, chuẩn bị cho sự phóng noãn và tương tác với tinh trùng. Kích thước noãn trưởng thành khoảng 120 um, thật sự đã đạt được trong quá trình phát triển của noãn. Các hormone hướng sinh dục được chế tiết bởi tuyến yên kiểm soát sự trưởng thành và phát triển của noãn. Ban đầu chúng tác động vào mô buồng trứng gây phóng noãn và sau đó kích thích các tế bào nang noãn giải phóng các yếu tố tác động trực tiếp vào màng bào tương noãn thúc đẩy trưởng thành tế bào.
Trong giai đoạn lớn lên, các tế bào hạt bao xung quanh phân chia nguyên phân và noãn bào tổng hợp 1 lớp áo glycoprotein dày 15 um với thành phần hoá học gồm 70% protein, 20% hexose, 3% sialic acid và 2% sulfat - được gọi là màng trong suốt của noãn. Đây là một cấu trúc phức tạp, có tác dụng bảo vệ noãn trong việc ngăn chặn đa tinh trùng thụ tinh và duy trì sự toàn vẹn của phôi trong giai đoạn sớm. Lớp ngoài cùng của màng có dạng lưới lỏng lẻo, gồm các dải sợi hạt, dày lên khi noãn bào tăng kích thước và có thể bị các enzyme, immunoglobulins và các virus nhỏ thấm qua. Các sợi nổi nhau liên tục ở mặt trong màng trong suốt ngắn và thẳng, với các cấu trúc tròn nhỏ tại các điểm giao nhau, và các sợi mảnh này hoà vào nhau khi thụ tinh. Các kênh bào tương đi qua màng trong suốt từ các tế bào hạt và kết thúc ở khe tiếp hợp. Chúng bị rút ra khi noãn bào bắt đầu trưởng thành, để lại những lỗ trống và sau đó được các chất gian bào làm đầy khi noãn bào trưởng thành hoàn toàn.
Có 3 loại glycoprotein chính được biết là ZP1, ZP2 và ZP3. Tinh trùng xâm nhập vào zona trong 2 - 15 phút và gắn vào zona qua trung gian ZP3 glycoprotein. Sự phân bố các carbohydrate và glycosaminoglycan thay đổi giữa noãn đang phát triển và noãn trưởng thành hoàn toàn. Màng trong suốt bị hoà tan khi phá vỡ các liên kết hoá trị II, do các protease hay dithiothreitol, hoặc các liên kết hóa trị bị phá vỡ bởi pH thấp và nhiệt độ cao. Lớp màng này trở nên không thể hoà tan sau khi thụ tinh do được che phủ 1 bởi lớp áo glycoproteins vòi tử cung
2.2 Sự hình thành và phát triển nang noãn
2.2.1 Sự hình thành nang noãn
Các noãn bào được bao quanh bởi những lớp tế bào sinh dưỡng đặc hiệu gọi là các tế bào nang. Noãn bào và các tế bào nang của nó kết hợp với nhau chặt chẽ. Các tế bào nang đảm trách việc chuyển các chất liệu dùng cho sự lớn lên của noãn bào và cung cấp các tín hiệu cho noãn bào trưởng thành. Lúc thai 18 - 20 tuần, tế bào vỏ dần dần bị thủng do các kênh mạch, và điều này đánh dấu sự bắt đầu hình thành nang noãn. Do sự xuất hiện các mạch máu hình ngón tay nhô vào trong vỏ, dẫn tới sự xuất hiện của các thừng sinh dục thứ phát. Do các mạch máu xâm nhập vào, chúng chia các khối tế bào vỏ trước đây thành các phần nhỏ hơn. Cùng với các mạch máu là các tế bào quanh mạch cũng đi vào. Những tế bào này bao quanh các noãn bào sơ cấp ở tiền kỳ giảm phân, tạo thành nang noãn nguyên thuỷ - gồm một noãn bào bị ngưng ở tiền kỳ giảm phân I, được phủ bên ngoài bởi một lớp đơn tế bào tiền hạt hình thoi, bao chung quanh là một màng đáy. Cuối cùng, tất cả các noãn bào được bao bọc theo kiểu thế này. Trung mô còn lại không tham gia vào sự hình thành nang noãn nguyên thuỷ hiện diện trong các khe giữa các nang, sẽ hình thành mô đệm buồng trứng nguyên thuỷ. Các tế bào hạt biệt hoá từ các tiền thân trung mô hoặc biểu mô. Quá trình này của sự phát triển các nang noãn nguyên thuỷ vẫn tiếp tục cho đến khi tất cả các noãn bào ở giai đoạn diplotene được bao bọc trong các nang, đôi khi kéo dài sau khi sinh.
.jpg)
Sau khi noãn bào được bao quanh bởi các tế bào tiền hạt, toàn bộ các nang noãn có thể trải qua các mức độ trưởng thành khác nhau. Sự hình thành nang noãn sơ cấp được đánh dấu bởi sự thay đổi lớp tế bào tiền hạt thành một lớp các tế bào hạt hình khối. Một số các nang hoàn thành giai đoạn lớn lên gọi là nang tiền khoang - nang thứ cấp là nang với các tế bào hạt tăng sinh hoàn chỉnh hơn và nhiều trong số này bị thoái hoá đi. Giai đoạn tiền khoang kéo dài 8 - 12 ngày và trong giai đoạn này có sự tích luỹ dịch trong nang bao quanh noãn bào với các tế bào hạt. Gonadotropins trong tuần hoàn chuyển các nang tiền khoang thành thành nang thật sự. Đỉnh LH sau đó gây ra sự tích luỹ dịch nhanh chóng lúc này gọi là giai đoạn tiền phóng noãn (nang De Graaf) - khoảng 36 giờ đặc trưng bởi một khoang chứa đầy dịch nang.
.jpg)
2.2.2 Ảnh hưởng của nội tiết tố sinh dục
Tuyến yên trước bắt đầu phát triển giữa tuần thứ 4 và thứ 5 của thai kỳ và tuần hoàn cửa vùng dưới đồi - tuyến yên đảm nhận chức năng vào tuần thứ 12. Nồng độ FSH của tuyến yên đạt đỉnh vào tuần 20 - 23 và nồng độ đỉnh trong tuần hoàn vào tuần 28. Nồng độ này cao hơn ở trong các bào thai giới nữ so với bào thai nam cho đến 6 tuần cuối của thai kỳ. Buồng trứng của các thai vô sọ do thiếu GnRH và chế tiết gonadotropins sẽ dẫn đến thiếu sự hình thành nang noãn trưởng thành và kích thước buồng trứng cũng nhỏ hơn khi thai đủ tháng, nhưng vẫn diễn ra sự giảm phân và phát triển của các nguyên thuỷ, rõ ràng là những điều này không phụ thuộc vào gonadotropins. nang noãn.
2.2.3 Kích thước nang noãn tương ứng với sự phát triển của noãn
Sự giảm phân không thể tiếp tục trong các noãn bào nằm trong các nang nguyên thuỷ hay nang sơ cấp (giai đoạn túi mầm kéo dài). Chỉ những noãn bào trong các nang De Graaf trưởng thành với kích thước bình thường mới có thể chuyển tiếp đến trung kỳ II. Đây là điểm quan trọng trong kích thích buồng trứng hỗ trợ sinh sản.
Việc gây trưởng thành noãn do đỉnh LH hoặc tiêm HCG, khởi phát một chuỗi các tương tác giữa các tế bào hạt, tế bào tia và noãn. Sự tương tác này được điều chỉnh thông qua việc chuyển hoá các loại tế bào này trong nang. Những chất trọng lượng phân tử thấp có thể khuếch tán giữa chúng, vì thế thông tin được chuyển nhanh chóng qua nang từ các thụ thể tế bào hạt.
Nang tiền phóng noãn trở nên không đáp ứng với LH do các thụ thể của chúng bị điều hoà giảm khi có đỉnh LH. Ngược lại đỉnh FSH làm tế bào hạt tổng hợp quá nhiều Hyaluronic Acid. Chondroitin sulfat và các glycosaminoglycans cũng được tổng hợp trong giai đoạn muộn của sự phát triển nang noãn. Noãn bào được bao quanh bởi một khối nhầy dày 1 cm hoặc hơn trong nang phóng noãn, có tác dụng bảo vệ noãn khi nang vỡ và khi noãn di chuyển vào trong vòi tử cung. Fibronectin và fibrinogen cũng kết hợp với hoạt động của tế bào hạt.
Sự nhạy cảm của các nang thay đổi có thể là một yếu tố điều hoà sự trưởng thành của noãn. Nó có thể liên quan đến sự thay thế các yếu tố ức chế bằng các yếu tố kích thích để khởi phát sự trưởng thành đáp ứng với đỉnh LH. Yếu tố ức chế trưởng thành có thể gồm AMP vòng, tác nhân ức chế trưởng thành noãn (OMI), hypoxanthine, dbcAMP. Các dấu hiệu trưởng thành gồm gonadotropin, yếu tố kích thích trưởng thành (MPF), calcium và EGF.
.jpg)
2.2.4 Sự phóng noãn
Sự phóng noãn là một quá trình giải phóng noãn có khả năng thụ tinh từ một nang trưởng thành. Thời gian phóng noãn rất thay đổi trong từng chu kỳ kinh, ngay cả trên cùng một người phụ nữ. Ước tính thời gian phóng noãn là 34 - 38 giờ sau đỉnh LH. Đỉnh LH khởi phát một chuỗi các biến cố:
Đỉnh LH kích thích sự tiếp tục phân chia giảm phân của noãn, hoàng thể hoá các tế bào hạt, tổng hợp progesterone và prostaglandin bên trong nang
Progesterone làm tăng hoạt động các men ly giải, cùng với prostaglandin làm vỡ thành nang.
Đỉnh FSH giữa chu kỳ góp phần làm chuyển plasminogen thành plasmin là men ly giải và đảm bảo đầy đủ thụ thể LH trên tế bào hạt để tạo giai đoạn hoàng thể bình thường.

.jpg)
2.3 Kết luận
Tóm lại, sự sản xuất giao tử ở nữ giới là một quá trình rất dài với nhiều giai đoạn được diễn ra theo một thứ tự nhất định. Sự hình thành và phát triển của noãn giúp tạo nên một noãn trưởng thành có khả năng thụ tinh. Đồng thời với nó là sự hình thành và phát triển các nang chứa noãn đảm bảo cho sự phát triển đầy đủ và phóng noãn hiệu quả.
So với sinh tinh, sự sinh noãn là rất ít ỏi trong mỗi chu kỳ. Nếu xảy ra bất thường trong giảm phân, tế bào noãn có thể chứa 22 hoặc 24 NST thay vì 23 NST như sinh lý. Trong trường hợp noãn bào chứa 24 NST được thụ tinh bởi tinh trùng bình thường (23 NST), sẽ tạo thành một hợp tử có 47 NST (hay còn gọi là thể ba nhiễm - trisomy). Trisomy 21 hay hội chứng Down là loại trisomy phổ biến nhất. Nếu một noãn có 22 NST được thụ tinh bởi một tinh trùng bình thường sẽ tạo ra một hợp tử có 45 NST, như hội chứng Turner (45,XO). Rối loạn bất cứ khấu nào trong các giai đoạn trên đều không thể sinh được noãn có khả năng thụ tinh. Nắm vững sinh lý quá trình sinh noãn giúp chúng ta có thể chủ động hơn trong công tác chẩn đoán và điều trị hiếm muộn.
2.4 Tài liệu tham khảo
1. Cao Ngọc Thành, Lê Minh Tâm (2017). Nội tiết phụ khoa và Y học sinh sản. Nhà xuất bản Đại học Huế.
2. Bruce M. Carlson (2019). Phôi người và Sinh học phát triển. Phiên bản thứ sáu. Elsevier.
3. Sadler T.W (2015). Phôi y học của Langman. Phiên bản thứ mười ba, Wolters Kluwer.
3 Sự sinh tinh
3.1 Giới thiệu
Tinh trùng người được Leeuwenhoek và Hamm quan sát lần đầu tiên vào năm 1677 bằng kính hiển vi cải tiến. Tuy vậy, họ không hiểu được vai trò của tinh trùng đối với quá trình sinh sản. Họ nghĩ rằng nó chứa bên trong một con người thu nhỏ và điều này được được xem như là lý thuyết tiên thành: con người thu nhỏ này sẽ lớn lên khi tinh trùng xâm nhập được vào trứng. Trước đó, vào năm 1675, Malpighi khi quan sát trứng gà chưa thụ tinh, đã gợi ý rằng quả trứng có chứa gà con thu nhỏ.
Lý thuyết tiền đề gây nhiều tranh cãi này đã bị bác bỏ vào năm 1775 khi Spallanzani chứng minh được rằng cả noãn và tinh trùng đều cần thiết cho sự hình thành nên một cơ thể mới. Vào năm 1841, Von Kolliker phát hiện ra rằng tinh trùng là sản phẩm cuối cùng của sự phân chia tế bào trong các tiểu quản sinh tinh của tinh hoàn.
Đến năm 1878, việc nghiên cứu nhiễm sắc thể (NST) của Flemming giúp ông ta có thể nhận ra NST có vai trò trong thụ tinh. Năm 1883, Van Beneden phát hiện rằng các tế bào mầm trưởng thành (giao tử) chứa số lượng NST giảm đi. Ông cũng mô tả sự phân bào giảm nhiễm, một quá trình làm giảm số lượng NST. Và cho đến 1956, người ta mới xác định được số lượng NST ở người là 46.
3.2 Sự sinh tinh
Không giống như sinh noãn, sự sinh tinh là một quá trình liên tục và có thể chia làm 3 giai đoạn: sự sản xuất các tế bào mầm (giao tử); sự biệt hoá chức năng để thụ tinh (giảm phân); và sự biệt hoá cấu trúc để hoạt hoá khả năng di động và xâm nhập noãn.
.jpg)
3.2.1 Sự tạo thành giao tử
Trong giai đoạn sớm của sự hình thành phôi, các tế bào mầm nguyên thuỷ di chuyển đến tuyến sinh dục đang phát triển. Để thực hiện sinh tinh, các tế bào này hiện diện ở các tiểu quản sinh tinh của tinh hoàn. Sau đó các tế bào mầm chưa trưởng thành, còn gọi là các tinh nguyên bào, sẽ phát triển từ các tế bào này thông qua nguyên phân.
Trong tiểu quản sinh tinh, có hai loại tế bào sinh dưỡng gồm tế bào cơ trơn, tế bào Sertoli và năm loại tế bào mầm gồm tinh nguyên bào (spermatogonia), tinh bào sơ cấp và tinh bào thứ cấp (spermatocyte), tinh tử (spermatid) và tinh trùng (spermatozoa). Các tinh nguyên bào nằm dọc theo bờ ngoài của các tiểu quản, tiếp xúc với các tế bào hỗ trợ, đó là các tế bào Sertoli. Các tế bào Sertoli tạo thành một lớp liên tục trên màng đáy của tiểu quản và gắn kết chắc chắn với nhau do ranh giới tế bào khít. Bằng cách này chúng tạo thành hàng rào tinh hoàn - máu về mặt miễn dịch.
Các tinh nguyên bào vẫn bị bất hoạt cho đến tuổi dậy thì. Từ khi dậy thì về sau, chúng liên tục phân chia bằng sự giảm phân để tăng số lượng tế bào và cung cấp nhiều tế bào mới một cách liên tục.
.jpg)
3.2.2 Sự biệt hoá về mặt chức năng
Khi các tế bào mầm nguyên thuỷ di chuyển hoàn toàn đến cầu sinh dục, chúng chiếm vị trí trung tâm của thừng sinh dục, nơi sẽ thành tiểu quản sinh tinh. Tại đây chúng được bao quanh bởi các tế bào Sertoli và một màng đáy. Tế bào sinh dục nam (gonocyte) không phân chia như ở nữ, chúng vẫn ở trạng thái bất hoạt cho đến ngay trước khi dậy thì.
3.2.2.1 Tinh nguyên bào (spermatogonia)
Các tế bào mầm nguyên thuỷ phân chia nguyên phân tạo ra các tỉnh nguyên bảo, là các tế bào lưỡng bội nằm ở lớp dưới cùng của tiểu quản, giữa các tế bào Sertoli và màng đáy. Các tính nguyên bào được chia làm 3 loại: A, trung gian và B. Tinh nguyên bào A được chia nhỏ thành supscripts 0, 1, 2, 3 hay 4. Tinh nguyên bào nguyên phân theo hai kiểu: phân chia ngẫu nhiên các tế bào mầm hay tinh nguyên bào AD (còn gọi là As) và kiểu phân chia đồng bộ của tỉnh nguyên bào A1, A2, A3, A4. Loại tinh nguyên bào trung gian và B được kết hợp với các khâu khác trong chu trình sinh tinh. Có thể thấy rõ sự khác biệt về mặt hình thái giữa các tinh nguyên bào A, trung gian và B nhưng giữa các subtype khác nhau của tinh nguyên bào A thì khó hơn.
Tinh nguyên bào A0 ở người trưởng thành chỉ phân chia khi các tinh nguyên bào khác bị phá huỷ, như tia X và khi tinh nguyên bào A4 phân chia sẽ làm tăng tinh nguyên bào trung gian hoặc một thế hệ tinh nguyên bào A1 mới. Nếu các biểu mô mầm không bị phá huỷ thì tốc độ phân chia của các tế bào mầm tăng lên để phục hồi lại tiểu quản sinh tinh.
.jpg)
3.2.2.2 Tinh bào sơ cấp (primary spermatocyte)
Ngay sau lần nguyên phân cuối cùng của tinh nguyên bào B, các tế bào còn gắn kết với nhau qua cầu nối bào tương sẽ nhân đôi bộ NST giống như để chuẩn bị cho nguyên phân một lần nữa. Tuy nhiên, lúc này nó bắt đầu một tiền kì giảm phân dài và sau đó là hai lần giảm phân để tạo ra 4 tinh tử đơn bội. Có thể ngay sau khi tổng hợp DNA (preleptotene phase) thì các tinh nguyên bào ở giai đoạn leptotene sẽ tách khỏi màng đáy, các tế bào Sertoli hai bên sẽ áp lại tạo nên phức hợp ranh giới thứ phát nằm giữa tinh nguyên bào và màng đáy. Khi đó tinh nguyên bào nằm phần giữa của tiểu quản và phần giữa tinh nguyên bào với lòng tiểu quản tách dần để tế bào mầm đi vào trong lòng và kết thúc quá trình giảm phân.
3.2.2.3 Sự giảm phân và tinh bào thứ cấp (secondary spermatocyte)
Ở giai đoạn hướng cực, các NST tương đồng tách ra, đánh dấu sự kết thúc lần giảm phân I. Nó xảy ra khá nhanh và mỗi tinh bào sơ cấp chia thành 2 tinh bào thứ cấp, mỗi tinh bào thứ cấp lúc này chứa bộ NST đơn bội ở dạng kép (1n2c). Thời gian tồn tại của tinh bào thứ cấp ngắn, nó nhanh chóng phân chia lần hai để chia đôi các NST dạng kép và thành hai nửa và thành hai tinh tử đơn bội.
3.2.3 Sự biệt hoá về cấu trúc
Các tinh tử trải qua sự biệt hoá về hình thái hình thành nên tinh trùng trưởng thành (spermatogenesis). Quá trình biệt hoá này được gọi là sự hình thành tinh trùng. Các tinh tử kéo dài ra và phát triển đuôi nhưng vẫn còn gắn với nhau và các tế bào Sertoli bên dưới bởi phương tiện là cầu nối tương bào. Kết quả của quá trình này là hình thành các tinh tử với đơn bội NST có nguồn gốc từ tế bào mẹ.
Cuối lần giảm phân thứ hai, tinh tử mới hình thành phối hợp phát triển với một thế hệ tinh tử hình thành trước đó một chu kỳ, nhưng vẫn còn phụ thuộc vào tế bào Sertoli. Sự phát triển của hai thế hệ này tiếp tục đồng bộ, các tinh tử chu kỳ trước được bao bọc bởi khoảng kẽ trên bề mặt của bào tương tế bào Sertoli còn những tinh tử mới hình thành nằm kẹp giữa các tế bào Sertoli như vị trí lúc chúng còn là các tinh nguyên bào và như khi giảm phân. Sự phát triển từ lúc giảm phân cho đến khi thành tinh trùng tách ra được gọi là sự sinh tinh. Tinh tử thời điểm ban đầu là tế bào tròn trông giống như tế bào bình thường với nhân rõ. Nhưng ngay sau đó có sự thay đổi, rõ nhất là sự cô đặc chromatin nhân. Thể Golgi tăng các hạt chứa dịch. Những hạt này hoà nhập vào nhau tạo nên một túi acrosome duy nhất. Rồi acrosome trải rộng ra bao quanh lấy bề mặt nhân.
Sau khi giảm phân, hai trung cực di chuyển đến phía đối diện với acrosome đang hình thành. Một trung cực bắt đầu phát triển về phía đầu xa để tạo thành đuôi, sau này với sự hình thành axoneme, còn trung cực kia tạo thành cổ hay đoạn nối để gắn đuôi tinh trùng vào nhân. Các sợi bên ngoài cô đặc phát triển từ bên ngoài thành các đội vi quản tương ứng của axoneme rồi sau đó tách ra, ngoại trừ đoạn cuối.
Lúc bắt đầu sự sinh tinh, các lưới nội bào là một hệ thống ba chiều các túi hình cầu và hình ống nối nhau bằng các tiểu quản hẹp. Hệ thống này trải rộng ra qua các cầu nối gian bào nổi tế bào với các tinh tử khác. Đến cuối giai đoạn sinh tinh, ER thoái triển và biến mất. Ty lạp thể của tinh tử đang phát triển tập hợp lại dần dần và tự nó bao quanh lấy đoạn giữa của tinh trùng và tạo thành một vòng xoắn đôi rất đặc trưng giữa cổ và annulus.
3.2.4 Sự giải phóng tinh trùng
Bước đầu tiên để giải phóng tinh trùng là phá vỡ phức hợp ống túi, có chức năng như một cái neo giữ tinh tử trong giai đoạn cuối phát triển. Việc tách phức hợp này cũng giúp loại đi một tỷ lệ lớn bảo tương của tinh tử (đến 70%). Phần bào tương còn lại của tỉnh tử vẫn gắn ở cổ tỉnh trùng. Khi tinh trùng di chuyển vào lòng tiểu quản, khối bào tương này không thay đổi theo tạo thành một dải bào tương mảnh dính giữa khối bào tương với tinh trùng và càng lúc càng dài ra. Ngay cả khi tinh trùng đã rời ra và đi vào lòng tiểu quản sinh tinh như một tế bào tự do, vẫn có thể còn một vết nhỏ dính ngay sau đầu tinh trùng được gọi là giọt bào tương. Phần bào tương còn lại phía tế bào Sertoli, được gọi là thể tồn dư, còn gắn với các thế dư của các tỉnh trùng khác qua cầu nối bào tương, sau đó sẽ bị thực bào bởi tế bào Sertoli. Chỉ khi sự biệt hoá kết thúc, các tinh trùng độc lập mới được giải phóng vào lòng các tiêu quản. Các tinh trùng cuối cùng đến được mào tinh, một ống xoắn nằm trong tinh hoàn, ở đó tinh trùng tiếp tục phát triển hơn nữa đến trưởng thành. Toàn bộ quá trình mất khoảng 64 ngày.
.jpg)
3.2.5 Khả năng sinh tinh
Tinh trùng đầu tiên được tạo ra từ tinh hoàn vào thời điểm dậy thì. Lúc này sự sản xuất androgen vẫn còn thấp hơn mức bình thường ở người trưởng thành. Tốc độ sản xuất tinh trùng ở mỗi tinh hoàn tăng lên theo độ lớn lên của tinh hoàn. Sự sinh tinh ở nam giới kéo dài liên tục suốt cuộc đời. Khi lớn tuổi, khả năng thoái hoá tăng lên, nhưng không có bằng chứng cho thấy có sự giảm kích thước tinh hoàn theo tuổi. Người ta ghi nhận khả năng sinh sản của đàn ông có thể kéo dài đến 90 tuổi.
3.3 Sự di chuyển tinh trùng
3.3.1 Sự di chuyển trong đường sinh dục nam
Tinh trùng mới hình thành nằm ở tiểu quản với đầu gắn vào các tế bào Sertoli. Dưới ảnh hưởng của nội tiết tố (testosterone) các tinh trùng được giải phóng vào lòng của tiểu quản và cuối cùng sẽ đến mào tinh. Mào tinh cung cấp dinh dưỡng cho tinh trùng và ảnh hưởng đến quá trình trưởng thành của chúng.
Tiếp theo mào tinh, tinh trùng đi vào ống dẫn tinh dài khoảng 45 cm. Sự di chuyển của tinh trùng nhờ vào những co thắt có tính nhu động của thành ống dẫn tinh. Khoảng cách tinh trùng phải di chuyển khoảng 6 - 7 m và mất 7 - 14 ngày.
3.3.2 Sự phóng tinh
Sự kích thích hệ thần kinh giao cảm dẫn đến sự co thắt nhu động các cơ mào tinh và ống dẫn tinh. Một lượng nhỏ dịch có chứa tinh trùng nằm ở mào tinh, ống dẫn tinh và bóng ống dẫn tinh sẽ được chuyển qua ống phóng tinh đến niệu đạo, tiền liệt tuyến. Tiếp đó túi tinh cũng chế tiết 2 - 3 ml dịch dính, kiềm tính qua ống phóng vào niệu đạo, tiền liệt tuyến.
Các cơ tiền liệt tuyến co thắt và thêm vào 1 - 2 ml dịch tiết của tiền liệt tuyến giống như nước có tính acid nhẹ. Niệu đạo tiền liệt tuyến tiếp tục vào niệu đạo. Sự co thắt các cơ quanh niệu đạo gây ra lực phóng tinh ra ngoài niệu đạo. Trong khi phóng tinh, bàng quang đóng lại để tránh phóng tinh ngược dòng.
Tinh dịch là một chất dịch đặc, bao gồm 2 thành phần gồm tinh trùng do tinh hoàn sản xuất và tinh tương được hình thành do 1/3 từ chất tiết tiền liệt tuyến, 2/3 từ chất tiết của túi tinh. Chất tiết tiền liệt tuyến chứa acid citric, acid phosphatase và Kẽm có pH thấp. Chất tiết từ túi tinh chứa Fructose và có pH cao. Thể tích bình thường thay đổi từ 2 - 5 ml, trong đó chỉ một tỷ lệ nhỏ do tinh trùng (10%) còn 90% do tinh tương.
Chức năng quan trọng nhất của tinh tương (với pH từ 7,2 - 7,8) là vận chuyển tinh trùng và làm tăng pH âm đạo (chức năng đệm) vì tinh trùng không thể di động nhanh ở pH dưới 6,2. Thể tích tinh tương ít sẽ không đủ khả năng đệm, dẫn đến sự bất động tinh trùng khi vào môi trường âm đạo.
Mỗi ngày hàng triệu tinh trùng được sản xuất, được lưu giữ ở mào tinh và được giải phóng từng đợt khi phóng tỉnh. Nếu sau 5 ngày không có sự phóng tinh xảy ra, chất lượng tinh trùng sẽ suy giảm với giả thuyết có một sự hồi tác từ mào tinh đến các tiểu quản, làm ức chế sản xuất tinh trùng. Một khả năng khác là sự tái hấp thu tinh trùng bởi mào tinh.
3.4 Nội tiết học sinh sản nam
Bên cạnh chức năng sản xuất giao tử, chức năng thứ hai của tinh toàn là sản xuất các nội tiết tố sinh dục. Mô tế bào giữa các tiểu quản chịu trách nhiệm sản xuất các hormone, androgen tinh hoàn. Mô liên kết này chứa các tế bào khe, được gọi là tế bào Leydig.
3.4.1 Sự sản xuất androgen
Hormone tinh hoàn quan trọng nhất là các steroids androgen, trong đó testosterone đóng vai trò quan trọng nhất, tiếp theo là androstenedione. Tinh hoàn cũng sản xuất một lượng nhỏ estrogen. Cả tinh hoàn và buồng trứng đều sản xuất estrogen và androgen. Tuy nhiên, tinh hoàn ưu thế sản xuất androgen và buồng trứng ưu thế sản xuất estrogen.
Testosterone được sản xuất bởi các tế bào Leydig. Các tế bào này phân tách với các tiểu quản sinh tinh bởi một khoang chứa dịch bạch huyết. Các tế bào Leydig tiết testosterone vào trong khoang này và theo dịch bạch huyết đến tiếp cận các tiểu quản.
Trong bào tương, testosterone được kết hợp với một protein kết hợp androgen (ABP) đặc biệt được chế tiết bởi tế bào Sertoli. Nồng độ testosterone trong huyết thanh ở nam giới trưởng thành từ 12 - 30 nmol/l (ở phụ nữ trưởng thành là 0,5 - 2 nmol/l) Nồng độ testosterone trong nước tiểu thay đổi giữa 100 đến 330 nmol/l/24 giờ (độ tuổi 20 - 40).
3.4.2 Chức năng của androgen
Androgen chịu trách nhiệm trong một số quá trình khác nhau:
- Kích thích sự sinh tinh.
- Khởi phát và duy trì các đặc tính sinh dục nam thứ phát từ lúc bắt đầu dậy thì.
- Kích thích sự phát triển đường sinh sản nam và duy trì hoạt động co giãn và chế tiết của nó.
- Duy trì biểu mô mầm của các tiểu quản trong tinh hoàn cùng với FSH.
- Hồi tác âm tính với thuỳ trước tuyến yên để ức chế giải phóng LH.
- Kích thích sự tổng hợp protein cho sự lớn lên và phát triển hệ xương.
- Ảnh hưởng đến hành vi tính dục và sự hoạt bát, năng nổ.
3.4.3 Điều hoà chức năng tinh hoàn
Ở nam giới, LH kích thích tế bào Leydig trong mô kẽ tăng tiết androgen, vì thế nó còn được gọi là hormone kích thích tế bào khe. Tầm quan trọng của FSH chưa rõ. Nó chỉ có ảnh hưởng rất ít hoặc không trực tiếp lên sự sản xuất androgen. Do có các thụ thể FSH trên các tế bào Sertoli, nên người ta cho rằng FSH kích thích sự sinh tinh trùng, nhưng sự ảnh hưởng này không vượt quá giai đoạn tinh bào sơ cấp. Sự sinh tinh hoàn chỉnh cần có testosterone.
Sự sản xuất LH và FSH được điều hoà bởi hormone giải phóng gonadotropin, sản xuất và chế tiết bởi vùng dưới đồi. Tất cả các hoạt động này được kiểm soát chặt chẽ bởi cơ chế hồi tác. Testosterone có ảnh hưởng ức chế lên vùng dưới đồi và vì thế tác động một cách trực tiếp đến sự giải phóng LH và FSH. Yếu tố điều hoà khác có nguồn gốc tinh hoàn, được gọi là inhibin, là một protein được sản xuất bởi các tế bào Sertoli.
.jpg)
3.5 Tinh trùng
3.5.1 Cấu trúc tinh trùng
3.5.1.1 Đầu tinh trùng
Hình dạng đầu của tinh trùng đặc trưng cho loài. Đầu tinh trùng người hình tròn và dẹt, kích thước dài 5 um, rộng 2,5 um và dày 1,5 um. Đầu tinh trùng của tất cả các loài động vật có vú chứa khối DNA-protein cô đặc cao được gọi là chromatin. Các chromatin được ổn định bởi những liên kết disulfide đầy năng lượng, thường tồn tại cho đến khi thụ tinh hoặc bị phá vỡ do các tác nhân hoá học. Khoảng hơn một nửa phía trước của đầu tinh trùng chứa thể cực đầu (acrosome), là một túi chứa enzym có màng bao bọc đóng vai trò quan trọng trong sự xâm nhập của tinh trùng vào trong trứng. Hình dạng của thể cực đầu cũng khác nhau tuỳ theo loài, nhưng nó thường nằm bao phía trước nhận, được bao bọc bởi màng bào tương với lượng bào tương khá nhỏ.
.jpg)
Vùng sau thể cực đầu là một phần quan trọng vì nó sẽ gắn vào và hoà nhập với noãn. Màng bào tương của vùng này được lót bên dưới bởi một lớp cô đặc dày gọi là tấm sau thể cực đầu (postacrosomal sheath). Một khoảng sáng và hẹp giữa lớp này và nhân kéo dài phía sau đến eo sau. Phần đầu gần sát đoạn giữa được biệt hoá cao tạo thành hố. Màng nhân của vùng này được bao phủ ở lớp ngoài bởi một lớp chất liệu cô đặc rất dày tạo thành đĩa nền.
3.5.1.2 Đuôi tinh trùng
Đuôi hay còn gọi là tiêm mao của tinh trùng được chia thành đoạn giữa, đoạn chính và đoạn cuối. Đoạn giữa kéo dài từ đầu tinh trùng đến hình xoắn ty lạp thể (mitochondrial helix). Phần gần đầu nhất được gọi là đoạn nối và gắn thể chuyển động vào nhân. Đây là một cấu trúc cực kỳ phức tạp. Ở phần đầu, một vùng động giống như một khớp lồi cầu được gọi là chỏm, gắn vào đĩa nền của nhân bởi nhiều sợi nhỏ mịn. Những sợi này bị phá vỡ hoặc hoà tan khi tách đầu tinh trùng khỏi đuôi. Từ phần chỏm trải dài lui sau là chín cột được phân chia riêng biệt, đến đầu xa được bao bọc bởi chín sợi bên ngoài cô đặc, nhờ đó nó được liên kết chắc chắn. Vùng cổ thường hơi mảnh và không có bào quan, ngoại trừ những nếp gấp của bao nhân và một hoặc hai ty lạp thể kéo dài. Những phần ty lạp thể còn lại xếp liên tiếp nhau kiểu tận- tận tạo thành hình xoắn chặt bao quanh các sợi bên ngoài cô đặc. Số vòng xoắn của hình xoắn ty lạp thể khác nhau nhiều giữa các loài, ở người khoảng vài vòng đến 15 vòng.
Ở trung tâm của đuôi là phức hợp sợi trục. Nó gồm hai vi quản ở trung tâm, bao quanh bởi chín đôi vi quản xếp cách quãng theo hình trụ. Mỗi đôi vi quản gồm 2 tiểu đơn vị: tiểu sợi A và B. Sợi A được gắn 2 cánh tay động hướng đến đôi vi quản bên cạnh. Những cánh tay này có hoạt động ATPase mạnh mẽ, có thể là nền tảng của sự di động của tinh trùng. Nếu không có chúng, tinh trùng sẽ bất động. Có các cầu nối nexin mảnh gắn kết các đôi vi quản với nhau. Các nang hoa hướng tâm gắn các đôi vi quản xung quanh với đôi vi quản trung tâm. Những vi quản này được tạo thành chủ yếu bởi một protein được gọi là tubulin, tương tự như protein được tìm thấy trong thoi vô sắc khi tế bào phân chia. Giữa sợi trục và hình xoắn ty lạp thể là 9 sợi bên ngoài cô đặc. Mỗi sợi bên ngoài như vậy chạy dọc sát bên ngoài đôi vi quản. Sợi bên ngoài giữa các loài khác nhau thì khác nhau về cấu trúc, kích thước, hình dạng và về bề dày. Gần đến đầu tận, các sợi bên ngoài gắn vào thành của đôi vi quản tương ứng.
.jpg)
Ở đoạn chính, phía sau hình xoắn ty lạp thể, các vi quản và sợi bên ngoài tiếp tục kéo dài, được bao quanh bởi tấm xơ gồm một số cung bao quanh chu vi của tinh trùng giữa hai cột dọc. Hai cột dọc này chạy dọc hai bên toàn bộ chiều dài của đuôi. Phần đầu tiên của tấm xơ gắn vào hai sợi bên ngoài (số 3 và số 8) và phần cuối kết thúc trước các sợi bên ngoài. Do đuôi tinh trùng thon nhọn, nên các cột nhỏ dần và các cung cũng mỏng dần. Tấm xơ, được cấu tạo bởi một protein duy nhất với trọng lượng phân tử khoảng 80.000, kết thúc cách chóp đuôi vài micron.
.jpg)
Ở đoạn cuối, phía sau đầu tận của tấm xơ, chứa 9+2 vi quản, được bao phủ bên ngoài bởi màng bào tương.
3.5.2 Chuyển hoá
Tinh trùng có thể sử dụng nhiều chất khác nhau, cả chuyển hoá kỵ khí và ái khí. Các đường đơn như Glucose, fructose hay mannose có thể bị phá vỡ nhờ men thuỷ phân glycolysis trở thành lactic acid, hoặc bị oxy hoá thành carbone dioxide và nước, tuỳ trường hợp. Tinh trùng cũng có thể oxy hoá nhiều chất hữu cơ khác, như lactate, pyruvat, các acid béo không no.
Nguồn năng lượng đầu tiên cho tinh trùng di động là nguồn các Adenine nucleotide: ATP, ADP và AMP. Tinh trùng cũng chứa nồng độ Carnitine cao, mà dạng oxy hoá của nó có thể đảm trách chức năng dự trữ chuyển hoá quan trọng. Tuy nhiên, nói một cách chính xác hơn là sự di động tinh trùng kiểm soát sự chuyển hoá. Nếu không di động, tinh trùng không có chuyển hoá cho dù có bất cứ sự điều khiển nào. Rất có thể tinh trùng tự tìm cho mình những chất ngoại sinh cần thiết trên đường di chuyển của nó trong đường sinh dục nam và nữ.
Tinh trùng khi được phóng tinh có khả năng tổng hợp kém hơn rất nhiều, so với lúc nó vừa rời khỏi tinh hoàn, vì có những thay đổi quan trọng xảy ra khi di chuyển qua mào tinh. Nếu tinh trùng được phóng ra tiếp xúc đột ngột với nhiệt độ thấp thì sẽ có phản ứng gọi là “sốc lạnh” ảnh hưởng đến khả năng sống và độ thẩm thấu của màng tế bào. Đây có thể là do ảnh hưởng của nhiệt độ lên lipid màng tinh trùng chứ không phải tổn thương chuyển hoá đặc hiệu.
3.5.3 Di động của tinh trùng
Protein góp phần lớn nhất để chuyển năng lượng hóa học thành năng lượng cơ học là dynein, với trọng lượng phân tử khoảng 500.000 và biểu hiện qua hoạt động ATPase. Dynein nằm ở cánh tay của các đôi vi quản. Tự thân các đội vi quản không thể co duỗi được, thật sự nó không thể rút ngắn lại được. Thay vào đó, các vi quản nằm cạnh nhau trượt lên nhau, đẩy tới bằng cách thay đổi tư thế của cánh tay động. Cánh tay động trên tiểu sợi A có hai tư thế: mở ra và khép lại. ATP được gắn vào cánh tay động ở tư thế khép. Cánh tay này mở ra khi ATP bị phá vỡ và gắn vào tiểu quản subfiber B của đội vi quản liền kề. Sau đó cánh tay này thay đổi góc gắn kết để tạo tư thế “dịch chuyển” khi các sản phẩm của sự thuỷ phân ATP được giải phóng. Việc tạo ra hay bổ sung ATP mới làm cho cánh tay động tách ra khỏi tiểu sợi B và trở về tư thế khép ban đầu.
.jpg)
Chuyển động của một nửa số vi quản của axoneme đồng bộ nhau, nhưng khác phá với nửa số vi quản còn lại. Vì thế, động tác trượt lên nhau này chuyển thành cử động uốn cong mặc dù người ta chưa biết nhiều là các vi quản phối hợp động tác trượt lên nhau như thế nào để tạo thành sóng nhu động để quẫy đuôi. Có thể cơ sở của cơ chế điều hoà này là sự phối hợp giữa đôi vi quản trung tâm và các nang hoa hướng tâm.
3.5.4 Phản ứng thể cực đầu
Ở vùng thấu quang của noãn có các thụ thể đặc hiệu loài. Nhờ vậy, chỉ tinh trùng người mới có thể kết hợp với noãn của người, tương tự như sự kết hợp của chìa khoá và ổ khoá. Sự kết hợp của tinh trùng vào vùng thấu quang nhờ phản ứng thế cực đầu. Sự hoà nhập này diễn ra giữa màng bào tương và màng thể cực đầu làm vỡ và giải phóng các thành phần thế cực đầu. Các thành phần này chứa nhiều enzyme gồm cả acrosin. Các tinh trùng cần những enzyme này để có thể xâm nhập vào vùng thấu quang. Khi đã tiếp cận đến noãn, sự hoà nhập xảy ra giữa màng bào tương của noãn và phần đầu của tinh trùng.
3.6 Kết luận
Tóm lại, sự sinh tinh là một quá trình hết sức phức tạp, điều này không có gì đáng ngạc nhiên khi chúng ta nhận thấy sự biệt hóa cao độ của tinh trùng. Với chất liệu di truyền bằng một nửa các tế bào sinh dưỡng - bộ nhiễm sắc thể đơn bội, cùng với quãng đường khá xa để đạt đến đích, trải qua nhiều hệ thống rào cản để thực hiện vai trò thụ tinh với noãn đã đòi hỏi tinh trùng phải đạt yêu cầu cao về cả hình thái, di truyền và sinh hoá học.
3.7 Tài liệu tham khảo
1. Bruce M. Carlson (2019). Human Embryology and Developmental Biology. Sixth Edition.
2. Cao Ngọc Thành, Lê Minh Tâm (2017). Nội tiết phụ khoa và Y học sinh sản. Nhà xuất bản Đại học Huế. Elsevier.
3. Nguyễn Thanh Tùng, Đặng Văn Phước, Đặng Công Thuận, Lê Minh Tâm, Nguyễn Vũ Quốc Huy (2020). Đánh giá sự thay đổi mô bệnh học tinh hoàn và quá trình sinh tinh ở chuột nhắt đực bị stress nhiệt mãn tính ở bìu. Tạp chí Hành vi Động vật và Khí tượng Sinh học. 8:174-180. https:// doi.org/ 10.31893/ jabb.20023.
4. John Hall, Michael Hall (2020). Guyton và Hall Giáo trình Sinh lý Y học, Elsevier. 5. Sadler T.W (2015). Phôi y học của Langman. Phiên bản thứ mười ba, Wolters Kluwer
4 Sự thụ tinh
4.1 Điều kiện giao tử trước thụ tinh
4.1.1 Điều kiện tinh trùng
Tinh trùng sau khi được hình thành trong lòng ống sinh tinh vẫn chưa có khả năng di chuyển hay thụ tinh được. Chúng phải trải qua giai đoạn trưởng thành thật sự sau một vài ngày khi di chuyển qua phần trên của mào tinh. Sự trưởng thành tại mào tinh giúp chuẩn bị cho tinh trùng có khả năng đáp ứng cao hơn với điều kiện của đường sinh dục nữ, với biểu hiện ở hầu hết các bào quan của tinh trùng. Sự thay đổi quan trọng liên quan đến khả năng thụ tinh của tinh trùng là sự phát triển khả năng di chuyển tiến tới trước. Chuyển động định hướng này là rất cần thiết cho tinh trùng khi xuyên qua lớp áo bao quanh noãn.
Bên cạnh đó, những thay đổi về chuyển hoá giúp cung cấp năng lượng cho hoạt động của đuôi tinh trùng. Trong khi di chuyển từ đoạn đầu, đến đoạn thân và đoạn đuôi của mào tinh, các kháng nguyên bề mặt tinh trùng cũng thay đổi, thêm và bớt các đại phân tử, thay đổi glycoprotein bề mặt như thay đổi sự gắn kết lectin của màng bào tương tinh trùng.
Những thay đổi khác về mặt cấu trúc nhằm tăng tính ổn định chromatin của nhân tinh trùng. Đó là do sự hình thành các cầu nối disulphide (-S-S-) giữa các protamine giàu cystein. Có sự hình thành liên kết chéo protein gắn SH giữa các sợi cô đặc, màng ty lạp thể và mảnh của đuôi, có ảnh hưởng lên khả năng bơi của tinh trùng trưởng thành.
Khi giao hợp, sự kích thích hệ thần kinh giao cảm dẫn đến sự co thắt nhu động các cơ mào tinh và ống dẫn tinh. Một lượng nhỏ dịch có chứa tinh trùng nằm ở mào tinh, ống dẫn tinh và bóng ống dẫn tinh sẽ được chuyển qua ống phóng tinh đến hiệu đạo tiền liệt tuyến. Tiếp đó túi tinh cũng chế tiết 2 - 3 ml dịch dính, kiềm tính qua ống phóng vào niệu đạo tiền liệt tuyến. Các cơ tiền liệt tuyến co thắt và thêm vào 1 - 2 mi dịch tiết của tiền liệt tuyến giống như nước có tính acid nhẹ, từ niệu đạo tiền liệt tuyến tiếp tục vào niệu đạo. Sự co thắt các cơ quanh niệu đạo gây ra lực phóng tinh ra ngoài niệu đạo. Trong khi phóng tinh, bàng quang đóng lại để tránh phóng tinh ngược dòng.
Dịch tiết của túi tinh chứa một số thành phần quan trọng: fructose (năng lượng chính cho tinh trùng sau khi xuất tinh), prostaglandin (kích thích sự co thắt cơ trơn đường sinh dục nam và nữ hỗ trợ sự di động của tỉnh trùng), hay fibrinogen (tiền chất của fibrin, làm đông tinh dịch sau xuất tinh). Dịch tiền liệt tuyến chứa acid citric, kẽm, magnesium và các fibrinogenase, aminopeptidase.
Tinh dịch có chứa tinh trùng khi vào đến âm đạo người nữ, dưới tác động pH acid của âm đạo (pH < 5), sẽ bị đông vón lại do sự hình thành fibrin để bảo vệ tinh trùng khỏi tác động môi trường acid và giữ tinh trùng ở vị trí gần tử cung tránh chảy ngược ra ngoài. Môi trường âm đạo sau đó bị kiềm hoá do khả năng đệm của tỉnh dịch với pH > 7,2. Sự ly giải bắt đầu xảy ra do tác động của fibrinogenase và giải phóng tỉnh trùng. Cơ chế này làm tăng tối đa số lượng tinh trùng đến được cổ tử cung.
Trong giai đoạn phóng noãn, do ảnh hưởng của estrogen, chất nhầy cổ tử cung nhiều và loãng, giúp cho sự di chuyển của tinh trùng qua dễ dàng hơn. Niêm mạc cổ tử cung có nhiều kẽ, giúp cho tinh trùng có thể khu trú tại các kẽ này. Từ đây tinh trùng có thể tiếp tục đi lên buồng tử cung và vào vòi tử cung. Như vậy, đoạn đường di chuyển của tinh trùng có thể chia thành 3 đoạn: qua cổ ngoài tử cung, giữ lại tại các kẽ của cổ tử cung và đi tiếp lên buồng tử cung. Chỉ khoảng 5 phút sau xuất tinh, người ta có thể tìm thấy tinh trùng trong vòi tử cung. Sự di chuyển của tinh trùng trong buồng tử cung và vòi tử cung nhờ vào sự co thắt cơ trơn, di chuyển của luồng dịch trong tử cung, các nếp gấp nội mạc và môi trường nội tiết vòi tử cung.
Quá trình trưởng thành hoàn chỉnh của tinh trùng cần một vài giờ tiếp xúc trong môi trường đường sinh dục nữ để có khả năng thụ tinh hoàn chỉnh. Đây là bước "khả năng hoá" tinh trùng. Khả năng hóa đặc hiệu theo loài. Ở người, tinh trùng có thể được khả năng hoá khi tiếp xúc với chất tiết đường sinh dục nữ. Thời gian cần thiết cho khả năng hoá khác nhau tuỳ loài, ở người là 6 giờ. Có hai thay đổi xảy ra: (1) tách các protein của tinh tương và mào tinh bao quanh tinh trùng, và (2) sau đó có sự thay đổi các glycoprotein trong màng bào tương tinh trùng. Khả năng hoá tinh trùng có thể xảy ra trong buồng tử cung, trong vòi tử cung hoặc trong ống nghiệm khi tinh trùng tiếp xúc với tế bào hạt - tế bào vành tia. Trong môi trường thí nghiệm, tinh trùng khi gắn chặt vào tế bào hạt trong 2 - 3 giờ sẽ được thay đổi các thành phần bề mặt do các tế bào này tiết ra glycosidase. Dịch nang noãn cũng có thể gây khả năng hoá tinh trùng trong môi trường thí nghiệm. Yếu tố vận động có trọng lượng phân tử thấp được tìm thấy trong dịch nang, buồng trứng, vòi trứng, tử cung có thể làm tăng khả năng chuyển hóa (và tăng di động) của tinh trùng bằng cách giảm ATP và tăng AMP vòng.
4.1.2 Điều kiện hoãn
Sự xuất hiện đỉnh LH huyết thanh trong mỗi chu kỳ kích thích những thay đổi cả ở nang noãn và ở noãn. Noãn bào sơ cấp đang ngưng trệ ở tiền kỳ giảm phân I - không có khả năng thụ tinh, sẽ được giải phóng, tiếp tục quá trình giảm phân. Khoảng 15 giờ sau đỉnh LH, nhân giai đoạn túi mầm sẽ bị phá vỡ, nhanh chóng hoàn tất lần giảm phân I, giải phóng thể cực thứ nhất, tiếp tục đi vào giảm phân II. Khoảng 35 giờ sau đỉnh LH, noãn thứ cấp ở trung kì giảm phân II sẽ bị ngưng trệ lần II. Noãn ở giai đoạn này, thường được chứa trong nang tiền phóng noãn (preovulated follicle), đã đủ khả năng thụ tinh và được phóng ra đi vào vòi trứng để gặp tinh trùng.
Noãn sau khi được phóng ra từ nang noãn sẽ được loa vòi trứng hứng lấy và chuyển vào bên trong. Noãn được dịch chuyển nhờ vào sự co thắt cơ trơn vòi tử sự chuyển động của các vi nhung mao, luồng dịch từ ổ phúc mạc. Tinh trùng và noãn gặp nhau ở 1/3 ngoài, tương ứng với đoạn bóng vòi tử cung.
Sự thụ tinh xảy ra là điều kiện để noãn tiếp tục hoàn tất quá trình giảm phân. Khả năng thụ tinh của noãn lúc này có được phụ thuộc vào khả năng tham gia vào các hiện tượng: (1) thu nhận đầu tinh trùng kiểu thực bào, (2) hiện tượng lan truyền các hạt vỏ, (3) tan màng nhân đầu tinh trùng và (4) phân rã chromatin của tinh trùng.
.jpg)
4.2 Quá trình thụ tinh
4.2.1 Phản ứng cực đầu
Phản ứng cực đầu là tình trạng thể cực đầu được “kích hoạt” hay bị phá vỡ để giải phóng các chất giúp tinh trùng xâm nhập vào noãn. Đây là một bước quan trọng cuối cùng để hoạt hóa tinh trùng tuyệt đối cần ion Ca2+ để xảy ra phản ứng Khi tăng độ pH trong môi trường khoảng 9 - 9,5 cũng có thể gây được phản ứng Như vậy, những thay đổi của phản ứng cực đầu cho phép các ion xâm nhập vào màng bào tương, thay đổi nồng độ ion Ca2+ tự do nội bào và kiềm hoá bào tương Nói tóm lại, chuẩn bị trước khi thụ tinh về phía noãn là noãn đang ở giai đoạn phân chia giảm phân trung kỳ II, còn về phía tinh trùng là sự tăng động, khả năng hóa và phản ứng cực đầu là các điều kiện cần thiết nhất định.
Khi tinh trùng tiếp xúc với noãn, phản ứng cực đầu xảy ra. Đầu tiên, màng bên ngoài acrosome và màng bào tương hoà nhập với nhau ở một số điểm ở nửa trước đầu tinh trùng. Sự hòa nhập giữa hai màng thế này tạo nên những khoảng trống (cổng ra) lớn dần và các chất chứa bên trong acrosome có thể thoát ra ngoài chứa ít nhất 12 enzyme khác nhau và acrosome nhanh chóng xẹp đi. Trong số những enzyme được biết của acrosome thì quan trọng nhất là hyaluronidase và một enzyme giống trypsin giàu năng lượng là acrosin, các enzyme khác như phosphatase arylsulfatase, collagenase, phosphorylase, phospholipase C, và ẞ-galactosidase.
Trong acrosome, men acrosin được dự trữ ở dạng pro-acrosin, chỉ khi có phản ứng cực đầu nó mới được giải phóng ở chuyển dạng hoạt động - acrosin. Hai enzyme này đều đóng vai trò quan trọng đối với sự xâm nhập của tinh trùng vào noãn. Hyaluronidase giúp làm tan chất acid hyaluronic giữa các cumulus bao quanh noãn. Sự hoạt hoá các thụ thể tinh trùng do ZP3 trên màng zona dẫn đến sự phosphoryl hóa protein và tăng Ca” nội bào. Các đường truyền tín hiệu dẫn đến sự phosphoryl hóa protein và phản ứng cực đầu gồm: (1) đường truyền adenylate cyclase - cAMP - proteinkinase A; (2) đường truyền phospholipase C - diacylglycerol - proteinkinase C; và (3) đường truyền guanylate cyclase - cGMP - protein G.
.jpg)
4.2.2 Sự tương tác giữa tinh trùng và màng thấu quang của noãn
4.2.2.1 Gắn kết tinh trùng - màng thấu quang
Sự gắn kết giữa bề mặt màng thấu quang với thành phần đặc hiệu trên màng bào tương của tinh trùng đã được khả năng hoá có thể là một sự liên kết phụ thuộc calcium và rất cần thiết cho sự xâm nhập của tinh trùng qua màng thấu quang. Sự gắn kết này cần có sự hiện diện receptor trên màng bào tương tỉnh trùng, dù có phản ứng cực đầu hay không. Vấn đề còn tồn tại là loại receptor nào trên màng bào tương và các yếu tố hỗ trợ nào trên màng thấu quang giúp cho sự gắn kết.
4.2.2.2 Di chuyển qua màng thấu quang
Người ta thừa nhận rằng bước gắn kết đặc hiệu như trên tạo cơ sở vững chắc cho tinh trùng chui vào màng thấu quang. Màng thể cực đầu đã phản ứng còn gắn lại ở bề mặt màng thấu quang. Tinh trùng thường chui vào màng thấu quang theo một đường hẹp và cong, dưới kính hiển vi điện tử, nó tạo với đường tiếp tuyến bề mặt noãn một góc khoảng 45°. Nhưng đôi khi tinh trùng có thể xâm nhập theo hướng thẳng đứng hay nằm ngang so với bề mặt noãn. Cùng với lực chuyển động cần thiết của đuôi, acrosin đóng một vai trò quan trọng trong sự xâm nhập của tinh trùng Thật sự acrosin được xem như là một “enzyme xâm nhập màng thấu quang” cho dù có thể cần phối hợp với những enzyme khác.
.jpg)
Tóm tắt các bước quan trọng trong sự xâm nhập của tinh trùng:
- Màng bên ngoài của thể cực đầu hoà vào với màng bào tương tinh trùng.
- Các hạt trong thể cực đầu vỡ ra giải phóng Lysine, các enzyme này “khoan” một đường qua màng ZP hoặc tạo sự thay đổi để các tiểu quản của acrosome có thể xâm nhập.
- Màng thể cực đầu bên trong kéo dài tạo thành các tiểu quản do sự đa trùng hợp các actin hay các sợi giống actin nằm ở vùng sau thể cực đầu.
- Khi đỉnh của các tiểu quản chạm đến màng bào tương noãn, hai màng này sẽ hòa nhập vào nhau.
4.2.3 Sự hòa nhập giao tử và hợp nhất tinh trùng vào noãn
Sau khi xuyên thủng và chui qua màng zona, tinh trùng nằm trong khoang quanh noãn và chờ một thời gian trước khi bám vào bề mặt màng bào tương noãn. Quan sát trên kính hiển vi điện tử thấy rõ ràng phần giữa đầu tinh trùng tương ứng với phần xích đạo hay vùng sau thể cực đầu tạo sự hoà nhập với noãn. Khi phần xích đạo tiếp xúc với màng bào tương noãn, thì phần màng bào tương này che lên phần xích đạo của tinh trùng tạo sự hoà nhập. Điều này cho thấy tầm quan trọng của vùng sau thế cực đầu được biệt hoá này. Tính ổn định đặc biệt của nó nhờ vào các liên kết protein cấu trúc giữa các màng song song, giúp nó không bị hoà tan trong phản ứng cực đầu.
Sự hoà nhập liên quan đến sự tương tác ban đầu của đầu tinh trùng với các vi nhung mao trên màng bào tương noãn. Sự hoà nhập này gây nên một số hiện tượng khác diễn ra trong vài phút gồm phản ứng vỏ, hoạt hoá noãn, hình thành và tổng xuất cực cầu II. Ngay sau khi hòa nhập vào noãn, màng nhân tinh trùng biến mất, bào tương noãn xuất hiện và chèn vào vùng sau nhân của đầu tinh trùng giúp nhấn chìm phần đầu tinh trùng. Phần màng bào tương che phủ vùng đầu phía sau và đuôi tinh trùng còn nằm lại trên bề mặt noãn.
4.2.4 Phản ứng vỏ
Noãn bào có sự tái phân bố các bào quan của bào tượng để chuẩn bị cho hợp tử chuyển thành phôi. Những bào quan như những túi nằm trong phức hợp Golgi, chứa enzyme và mucopolysaccharides. Trong quá trình hoạt hoá, các hạt vỡ ra giải phóng các chất chứa bên trong vào trong khoang quanh noãn. Có hai kết quả ngay tức thì của hiện tượng lan truyền ngoại bào của các hạt vỏ gồm tăng thể tích khoang quanh noãn và màng zona chuyển thành cấu trúc dày cứng để bảo vệ. Phản ứng vỏ đảm bảo sự thụ tinh bình thường, một tinh trùng thụ tinh với một noãn, tránh hiện tượng đa tinh trùng thụ tinh.
Hiện tượng lan truyền ngoại bào bắt đầu khoảng 10 giây sau khi tinh trùng xâm nhập vào màng zona. Sự trì hoãn này gọi là giai đoạn tiềm tàng. Các hạt tại vùng cạnh vị trí tinh trùng xâm nhập bị phá vỡ đầu tiên và tạo nên một làn sóng lan rộng chậm xung quanh bề mặt noãn bào. Các mucopolysaccharides được giải phóng từ các hạt vào trong khoang quanh noãn gây nên sự tràn dịch nhanh chóng làm căng phồng màng thấu quang và giãn cách 1 - 20 km so với bề mặt noãn bào.
.jpg)
Diễn tiến dạng sóng của sự lan truyền ngoại bào cùng với sự nâng lên của màng zona mất khoảng 20 giây và lan rộng quanh bề mặt noãn bào. Hơn 5 phút sau, khoang quanh noãn tiếp tục tăng lên và thấu quang dày hơn, cứng hơn.
Mặc dù người ta chưa rõ phản ứng vỏ bắt đầu như thế nào hay lan truyền như thế nào xung quanh noãn bào, nhưng một yếu tố quan trọng là sự tăng Ca2+ trong bào tương; người ta có thể gây ra hiện tượng lan truyền ngoại bào các hạt vỏ bằng cách tiêm Ca2+ vào bào tương noãn và nó cũng được ngăn chặn bằng cách dùng tác nhân chelating hoá Ca2+ (những phân tử kết hợp Ca tự do). Ngay sau khi phát hiện thụ tinh, có sự giải phóng một lượng Ca từ nơi lưu trữ nội bào, gây nên sự tăng Ca2+ từ 0,1 - 10 um. Trong vòng vài phút, Ca2+ trở về vị trí nghỉ. Vùng tăng Ca2+ bắt đầu tại điểm hoà nhập tinh trùng - noãn và truyền ngang qua noãn với những sóng tương tự nhỏ hơn tiếp tục nhiều phút sau thụ tinh. Cơ chế của sự lan truyền sóng chưa rõ. Những sóng Ca” này tái diễn và kéo dài trong khoảng thời gian 3 phút.
4.3 Hoạt hóa noãn và hợp nhất tiền nhân
4.3.1 Hoạt hoá noãn
Sự di động của tinh trùng trong khoang quanh noãn nhanh chóng bị ức chế do hiện tượng hoà nhập tinh trùng. Lúc này có sự thay đổi điện tích bề mặt noãn, phản ứng vỏ xảy ra ngăn hiện tượng đa thụ tinh. Sự hoà nhập kích thích hình thành thoi vô sắc giai đoạn trung kỳ giảm phân II của noãn thứ cấp, các NST kép tách nhau ra ở tâm động đi về hai cực tế bào. Một nửa bộ NST sẽ được tống xuất ra ngoài noãn bào vào trong khoang quanh noãn theo thể cực thứ hai, còn lại noãn bào với bộ NST đơn bội.
Noãn trưởng thành (MII) chứa MPF ổn định và được hoạt hoá (M-phase Promoting Factor - MPF - yếu tố thúc đẩy phase M), một yếu tố kiểm soát chu kỳ tế bào cần thiết, là một protein-serin/threonine kinase, gồm có cdc2 và cyclin B; và được ổn định bởi c-mos proto-oncogene là một yếu tố ổn định tế bào. Sự hoạt hoá và bất hoạt MPF xảy ra tuỳ vào sự phosphoryl hóa hay dephosphoryl hóa, phối hợp với các phân tử chu kỳ tế bào chính. Noãn thu được sau chọc hút trong IVF tăng dần tỷ lệ MII sau khi cấy 4-6 tiếng và tình trạng hoạt hoá noãn duy trì cho đến khi tinh trùng xâm nhập. MPF bị bất hoạt khi đầu tinh trùng bị rã và hình thành màng nhân bao quanh tiền nhân đực.
4.3.2 Sự hình thành và hợp nhất tiền nhân
Khi đầu tinh trùng đi vào bào tương noãn, ngay lập tức màng nhân bị tan ra và chất nhân bị rã. Màng nhân tinh trùng bị tan ra do sự hoà lẫn lớp trong và lớp ngoài. Sự phồng lên của chromatin tinh trùng cô đặc cần thiết phá vỡ các liên kết chéo disulphide, nhờ tác dụng của yếu tố phát triển tiền nhân đực. Nhân tinh trùng lớn lên thành một mạng lưới các dải nucleoprotein, các protamine giàu Cystein có hàm lượng cao Arginine biến mất khỏi vị trí kết hợp với DNA tinh trùng. Sự trải rộng chromatin tinh trùng để hình thành tiền nhân có thể quan sát được nhờ vào sự hình thành màng nhân và sau đó là sự hiện diện của các hạt nhân con cô đặc. Sự hình thành tiền nhân đực phải được diễn ra trong điều kiện đặc hiệu của bào tương noãn giai đoạn trưởng thành, không thể xảy ra khi noãn chưa trưởng thành. Tiền nhân đực tích cực hơn trong sự hình thành tiền nhân. Sự sao chép DNA diễn ra trong thời gian từ 5 - 8 giờ sau khi tinh trùng xâm nhập.
.jpg)
Sự hình thành tiền nhân cái tương đối ít phức tạp hơn do chromatin của noãn không bị cô đặc cao như ở tinh trùng và không cần trao đổi protein nhân. Phần chromatin còn lại sau khi thể cực thứ hai bị tống xuất được trải rộng và nó chịu sự thay đổi tương ứng để trở thành tiền nhân cái với màng nhân bọc ở ngoài, tương tự như ở tế bào sinh dưỡng bình thường. Tuy nhiên, đến giai đoạn cuối, siêu cấu trúc hai tiền nhân tương tự nhau và không còn có thể phân biệt tiền nhân đực và tiền nhân cái. Tiền nhân đực và tiền nhân cái được hình thành lúc này di chuyển đến gần nhau rồi cùng di chuyển đến trung tâm của noãn. NST của tinh trùng và noãn trộn lẫn vào nhau, tạo thành trung kỳ của lần nguyên phân đầu tiên.
5 Tài liệu tham khảo
- Lê Minh Tâm, Cao Ngọc Thành, Trần Thị Như Quỳnh, Nguyễn Đắc Nguyên. Các vấn đề trọng yếu trong hỗ trợ sinh sản.
- Bruce M. Carlson (2019). Phôi người và Sinh học phát triển. Phiên bản thứ sáu. Elsevier.
- Cao Ngọc Thành, Lê Minh Tâm (2017). Nội tiết phụ khoa và Y học sinh sản. Nhà xuất bản Đại học Huế.
- Linda M. Ernst, Eduardo D. Ruchelli Chrystalle Katte Carreon Dale S. Huff (2019). Atlas màu về mô học của thai nhi và trẻ sơ sinh. Springer Nature Thụy Sĩ AG
