Các giai đoạn phát triển phôi và quá trình làm tổ của phôi người
1 Sự phát triển của phôi trước làm tổ
Nuôi cấy phôi là giai đoạn quan trọng nhất trong chu kỳ điều trị hỗ trợ sinh sản, Trong đó, vai trò của các yếu tố như kỹ năng và kinh nghiệm của chuyên viên phối học, điều kiện labo, tủ cấy và hệ môi trường nuôi cấy đều có ý nghĩa quan trọng. Chi tiết về sự phát triển của phôi trong quá trình nuôi cấy được mô tả trong tập 2 - Quy trình dành cho chuyên viên phôi học. Nội dung bài này dành cho bác sĩ lâm sàng hỗ trợ sinh sản nhằm mục đích nắm được các mốc sinh lý cơ bản để có thể ra quyết định điều trị.
1.1 Sự phát triển phôi qua các giai đoạn
1.1.1 Phôi giai đoạn phân cắt
Sau khi hai tiền nhân tiếp cận và hoà nhập vào nhau, hợp tử ở vào giai đoạn trung kỳ của lần phân chia đầu tiên. Quan sát dưới kính hiển vi vào thời điểm 18 giờ sau thụ tinh để tìm hai tiền nhân (thường kèm hai thể cực) là dấu hiệu chắc chắn thụ tinh đã xảy ra. Sự phân chia lần I kết thúc sau thụ tinh 24 giờ tạo thành phôi 2 tế bào, là chu kỳ kéo dài nhất, các chu kỳ sau chỉ khoảng 18 giờ. Những phân chia này giống như nguyên phân của tế bào bình thường, các tế bào con được tạo ra gọi là các phôi bào. Phân chia lần II kết thúc sau thụ tinh 40 giờ, tạo thành phôi với 4 phôi bào kích thước tương đương nhau. Vào ngày 3, phôi chứa 6 - 12 tế bào và ngày 4 gồm từ 16 - 32 tế bào.
.jpg)
Tuy nhiên, giai đoạn phân tách ngược lại với sự sinh noãn khi nó chỉ nhân đôi rồi phân chia NST và phân tách tế bào mà không có sự lớn lên (sự sinh noãn chỉ lớn lên chứ không nhân đôi và không phân chia). Đối với các tế bào sinh dưỡng, sau mỗi lần nguyên phân, nó phải tổng hợp bào tương để tăng kích thước trước khi có thể tiếp tục lần nguyên phân tiếp theo, để duy trì kích thước ổn định. Các phôi bào con được sinh với kích thước chỉ bằng một nửa tế bào ban đầu và chúng vẫn tiếp tục phân chia nhỏ hơn và nhỏ hơn. Đây là điểm khác biệt rất quan trọng của giai đoạn phân tách.
Càng về sau, sự khác biệt giữa các phôi bào càng tăng lên do sự phân chia các thành phần bào tương không đồng đều hoặc do những thay đổi diễn ra trong bản thân phôi bào khi phát triển. Nhân của mỗi phôi bào sẽ được định hướng theo môi trường bào tương khác nhau và vì thế ảnh hưởng lên hoạt động genome cũng khác nhau. Kết quả là mỗi phôi bào sẽ thiết lập cho riêng nó một chương trình phát triển và sẽ tăng dòng tế bào đặc hiệu. Những lần phân chia đầu tiên thường đồng bộ, nhưng những lần sau thì không. Các phôi bào được tổ chức thành từng nhóm hay từng lớp và mỗi nhóm này có tốc độ phân tách đặc trưng riêng. Đây là những cơ sở cho sự hình thành và biệt hoá cơ quan sau này như thần kinh, cơ, xương...
Kích thước toàn bộ của phôi không thay đổi gì trong giai đoạn phân tách mà vẫn giữ nguyên hình dạng của màng zona. Nuôi dưỡng cho phôi giai đoạn này do tự bản thân các phức hợp tế bào hạt - vành tia bao quanh nó và dịch tiết của biểu mô vòi trứng. Protein dịch tiết vòi trứng biểu hiện dạng điện chuyển như trong huyết thanh, kết hợp với B-glycoprotein.
1.1.2 Giai đoạn phôi dâu
Sau một vài lần phân chia, phôi chứa từ 16 - 32 tế bào có hình dáng như trái dâu nên gọi là phôi dâu. Giai đoạn này phôi đã xuống đến buồng tử cung. Sau lần phân chia thứ ba, trong phôi diễn ra quá trình kết đặc tế bào. Hiện tượng kết đặc tế bào xảy ra vào khoảng ngày thứ 3 - 4 làm cho các tế bào áp sát vào nhau, tăng tối đa sự tiếp xúc giữa các tế bào, khoảng gian bào giảm đi, các đường viền quanh tế bào cũng mờ đi và ranh giới giữa các tế bào lúc này trở nên khó phân biệt. Giai đoạn này có gia tăng đáng kể sự tổng hợp protein và DNA. Quá trình kết đặc tế bào phụ thuộc vào calci và liên quan đến khung tế bào và phân tử uvomorulin kết dính tế bào (còn gọi là yếu tố kết dính cadherin). Lúc này các phôi bào trở nên phân cực hóa rất cao, tạo điều kiện cho sự biệt hoá tế bào, các phôi bào chia thành 2 nhóm: nằm ở trung tâm là khối tế bào trong (inner cell mass - ICM), và một lớp bao quanh là khối tế bào ngoài. Do sự kết dính khác nhau, mặt ngoài các tế bào trở nên lồi và mặt trong trở thành lõm. Trong quá trình phân chia, các tế bào bên ngoài có được các đặc tính của biểu mô, hình thành các cầu nối gian bào, tạo nên ranh giới chặt chẽ rất khó tách rời. Cấu trúc này giúp ngăn cách lớp tế bào bên trong tiếp xúc với môi trường bên ngoài.
Quá trình kết đặc tế bào rất quan trọng trong sự biệt hoá khối tế bào trong và tế bào lá nuôi, quyết định đến sự hình thành của phôi thai. Nếu hiện tượng này bị ức chế do kháng thể kháng uvomorulin, thì phôi vẫn có thể phát triển thành phôi nang nhưng nó chỉ gồm một túi các tế bào lá nuôi mà không có khối tế bào trong và vì thế không thể phát triển thành phôi sống được. Các nguyên bào phôi giai đoạn này đều có hình tròn, thống nhất về mặt hình thái, sinh hoá, tiềm năng phát triển. Tất cả các nguyên bào phôi đều có thể tham gia vào sự hình thành tổ chức phôi, chúng có tính toàn năng. Tuy nhiên về sau, các đặc điểm này của phôi bào thay đổi đi. Màng thấu quang trong giai đoạn phôi dâu vẫn nguyên vẹn tạo điều kiện cho sự kết đặc tế bào và ngăn hai phôi (nếu có) nhập vào nhau. Nếu lớp tế bào trong phân chia ở giai đoạn sớm này có thể phát triển thành song thai cùng hợp tử.
1.1.3 Giai đoạn phôi nang
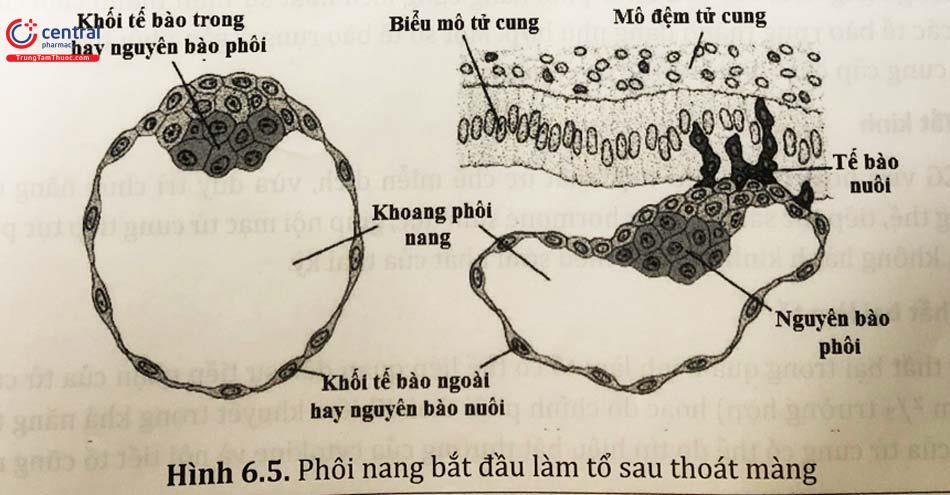
Phôi dâu nằm trong lòng tử cung được vùi trong niêm dịch của tử cung. Với sự hấp thụ của phôi, chất dịch này thấm qua màng thấu quang, vào giữa các phôi bào tạo thành những túi dịch nhỏ. Các túi dịch tăng dần kích thước, lớn dần và sáp nhập vào nhau tạo nên một khoang lớn là khoang phôi nang. Do ranh giới giữa các tế bào bên ngoài chặt, dịch tiếp tục đi vào chủ yếu nằm giữa các tế bào của khối tế bào bên trong. Khối tế bào bên trong bị chèn ép tạo thành khối cô đặc và bị đẩy về một phía của khoang phôi, chính là mầm phôi và sẽ phát triển thành cơ thể phôi thai. Khối tế bào bên ngoài trở thành một lớp biểu mô mỏng sẽ phát triển thành lá nuôi để tạo nên bánh nhau và các phần phụ của thai. Nơi mầm phôi nhô vào bên trong lòng khoang phôi nang gọi là cực phôi, đối diện với cực phôi là cực đối phôi. Các tế bào của phôi nang khác biệt nhiều so với các tế bào trong những lần phân chia đầu. Tuy nhiên sự khác biệt giữa lá nuôi và mầm phôi lúc này chưa đáng kể.
.jpg)
1.2 Sự phân chia tế bào phôi
Không giống như các tế bào cơ thể khoẻ mạnh bình thường, phôi bào không cần các phân tử tín hiệu đặc trưng để khởi động sự phân chia. Liên quan đến sự điều hoà phân chia tế bào, phôi tự điều hoà quá trình này nhờ vào đồng hồ nội bào. Các giai đoạn phân chia như sau:
Phase G1: diễn ra sự tổng hợp RNA (sao mã) và tổng hợp protein. Trong suốt giai đoạn này, tế bào không phân chia. Thời gian kéo dài khác nhau của giai đoạn này sẽ quyết định tốc độ phân chia tế bào. Sự tổng hợp DNA xảy ra ở phase S.
Phase G2: là phase chuyển tiếp từ phase S sang phase M. Phase M là giai đoạn nguyên phân, là lúc bắt đầu cô đặc và lúc kết thúc phân rã NST. Cô đặc là một quá trình cần thiết cho sự hình thành cấu trúc NST đặc hiệu của DNA. Sau khi hoàn tất phân rã (decondensation) NST, tế bào trở lại phase G1. Chu kỳ tế bào được điều hoà ở 3 điểm, không phụ thuộc vào sự phân chia nguyên phân hay giảm phân: (1) quá trình khởi động ở phase G1, (2) trước khi bắt đầu phase M, (3) giai đoạn trung kỳ. Phosphoprotein pp34 đóng một vai trò thiết yếu trong 3 quá trình tế bào này, kể cả tế bào cơ thể bình thường và tế bào phôi.
Trước khi bắt đầu nguyên phân, pp34 phóng thích nhóm phosphorus và kết hợp với cyclin, một protein chính yếu khác trong điều hoà chu kỳ tế bào. Toàn bộ phức hợp pp34-cyclin được gọi là yếu tố thúc đẩy trưởng thành MPF. Khi cyclin trong phức hợp này được phosphoryl hoá sẽ hoạt hoá enzyme và đảm bảo được sự cô đặc nhiễm sắc thể để bắt đầu phase M. Tác động kinase của phức hợp MPF cần thiết cho sự cô đặc nhiễm sắc thể. Như vậy, protein trong các NST chịu sự phosphoryl hoá dưới ảnh hưởng của PME. MPF không ổn định, nó hoạt hóa một protease làm phá vỡ đơn vị cyclin trong phức hợp MPF-cyclin, làm bất hoạt MPF. Sau đó, các tế bào đi vào trung kỳ nguyên phân và NST phân rã. Trong phase G1, một cyclin đặc hiệu với G1 kết hợp với pp34 tạo ra phase S. Sau khi phức hợp này phân ly, chu kỳ tế bào lại có thể bắt đầu.
Bình thường, sự phân chia bào tương và phân chia nhân diễn ra đồng đều, NST sẽ được chia đều cho hai tế bào con. Khi sự phân chia này không đồng đều sẽ tạo ra các phôi bào có bộ NST bất thường, phôi bào có nhiều nhân hoặc không có nhân. Các bào tương không nhân tiếp tục phân chia cho đến khi chỉ còn lại một mảnh bào tương không nhân.
1.3 Biểu hiện gen và sự tổng hợp protein của phôi
Trước khi thụ tinh, noãn trưởng thành đã có cấu tạo sinh hoá hoàn chỉnh cho sự tổng hợp protein. Trong giai đoạn lớn lên, noãn đã chứa đủ các ribosome (bào quan tổng hợp protein), RNA ribosome (rRNA), RNA thông tin (mRNA) và RNA vận chuyển (tRNA). Trong chu kỳ tế bào đầu tiên sau thụ tinh, protein được tổng hợp, điều chỉnh và bị phá vỡ vào các thời điểm cụ thể. Quá trình này vẫn tiếp tục trong thực nghiệm kể cả khi lấy nhân ra khỏi phôi và khi có các tác nhân ức chế đặc hiệu sự tổng hợp RNA. Điều này cho thấy, trong những chu kỳ tế bào đầu tiên, sự chuyển hoá và tổng hợp protein hoàn toàn chịu sự kiểm soát của thông tin mà noãn đã nhận được từ mẹ trong quá trình phát triển và trưởng thành.
Các thông tin di truyền của phôi, kể cả phần xuất phát từ bố chỉ được dùng ở giai đoạn sau. Điều này xảy ra sau khi hoạt hoá genome của phôi, gọi là sự chuyển tiếp thông tin từ mẹ sang hợp tử (Maternal to zygotic transition - MZT). Mặc dù, những kỹ thuật sinh hoá và sinh học phân tử nhằm phát hiện MZT chưa được thực hiện trên phôi người, nhưng dựa vào những thay đổi hình thái nhân chứng tỏ sự chuyển thông tin di truyền từ mẹ sang phôi diễn ra vào khoảng giai đoạn 8 tế bào.
Nghiên cứu dạng protein khi thông tin mẹ truyền sang phôi cho thấy, một nhóm protein có trọng lượng phân tử giống nhau từ 65.000 - 70.000 được tạo ra đầu tiên. Nhóm này gọi là protein yêu cầu sao mã (TRPs) và biến mất sau giai đoạn 8 tế bào. Khi sự tổng hợp DNA và phân chia tế bào trong phôi đã bị ức chế hoàn toàn thì lúc này TRP vẫn còn hiện diện. Điều này cho thấy sự giải mã được khởi đầu nhờ đồng hồ sinh học và chạy liên tục, không phụ thuộc vào sự tổng hợp DNA và phân chia tế bào.
Sự truyền thông tin từ mẹ sang phôi có ý nghĩa quan trọng về mặt sinh hoá và thực hành khi thực hiện nuôi cấy phôi. Trong thực hành, giai đoạn MZT diễn ra trùng với giai đoạn phôi ngừng phân chia nếu điều kiện môi trường thiếu sót. Hiện nay, người ta vẫn chưa hiểu điều kiện môi trường ảnh hưởng như thế nào đến MZT và liệu có liên quan đến các phân tử tín hiệu đặc trưng hay không. Như vậy, sự phát triển của phôi trước khi làm tổ chỉ diễn ra trong tuần đầu tiên sau thụ tinh. Hợp tử ban đầu trải qua giai đoạn phân chia làm tăng số lượng các phôi bào. Phôi dâu hình thành khoảng ngày 3 - 4 với đặc trưng hai khối tế bào bên trong và bên ngoài. Sự hình thành khoang phôi giữa các tế bào trong tạo nên phôi nang cùng với sự biệt hoá khối tế bào bên trong thành mầm phôi và biệt hoá khối tế bào bên ngoài thành lá nuôi. Chúng là cơ sở ban đầu cho sự hình thành cơ thể phôi cũng như bánh nhau và các phần phụ sau này. Phôi nang sẽ bắt đầu làm tổ ở nội mạc tử cung sau khi thoát khỏi màng thấu quang. Song song với những thay đổi về mặt hình thái là sự biến đổi mạnh mẽ về chuyển hoá, sinh tổng hợp protein và chuyển đổi gen. Vì thế, dù chỉ diễn ra trong thời gian ngắn, nhưng những thay đổi có tính quyết định của giai đoạn này làm cho nó đóng một vai trò vô cùng quan trọng trong sự sinh sản người.
1.4 Tài liệu tham khảo
1. Bruce M. Carlson (2019). Phôi người và Sinh học phát triển. Phiên bản thứ sáu. Elsevier. hạn
2. Cao Ngọc Thành, Lê Minh Tâm (2017). Nội tiết phụ khoa và Y học sinh sản. Nhà xuất bản Đại học Huế.
3. Lê Minh Tâm, Nguyễn Văn Trung (2015). Yếu tố phân chia phôi sớm trong dự đoán sự phát triển của phôi thụ tinh ống nghiệm. Tạp chí y học thực hành. Số 3 (954): 36-39.
4. Lê Minh Tâm, Nguyễn Văn Trung Nguyễn Thị Thái Thanh, Nguyễn Thị Tâm An, Cao Ngọc Thành, Lê Đình Dương, Alan H. DeCherney, M. Blake Evans, Micah J Hill (2018). Rã đông chuyển phôi sau nuôi cấy qua đêm so với nuôi cấy 2 giờ Một thử nghiệm triển vọng. Trình bày miệng. Bài viết 70. Đại hội lần thứ 8 của Sáng kiến Châu Á Thái Bình Dương về Sinh sản (ASPIRE 2018), Đài Bắc, Đài Loan từ ngày 12 - 15 tháng 4 năm 2018.
5. Linda M. Ernst, Eduardo D. Ruchelli Chrystalle Katte Carreon Dale S. Huff (2019). Atlas màu về mô học của thai nhi và trẻ sơ sinh. Springer Nature Thụy Sĩ AG.
6. Minh Tam Le, Trung Van Nguyen, Tung Thanh Nguyen, Thai Thanh Thi Nguyen, Tam An Thi Nguyen, Quoc Huy Vu Nguyen, Ngoc Thanh Cao (2019). Hội chứng buồng trứng đa nang có ảnh hưởng đến hình thái động học hoặc những bất thường trong quá trình phát triển phôi sớm không? Tạp chí Sản Phụ khoa và Sinh học Sinh sản Châu Âu: X3 100045. https:// doi.org/ 10.1016/ j. eurox.2019.100045
7. Sadler T.W (2015). Phôi y học của Langman. Phiên bản thứ mười ba, Wolters Kluwer
2 Sự làm tổ của phôi người
Quá trình mang thai thành công nhờ vào một loạt các sự kiện quan trọng bao gồm phôi làm tổ vào niêm mạc tử cung, màng rụng hoá, phát triển của nhau thai và sinh con. Mỗi sự kiện này đều rất quan trọng để đảm bảo thai kỳ diễn tiến. Quá trình làm tổ đòi hỏi một sự đối thoại mật thiết giữa phôi thai và nội mạc tử cung với khả năng tiếp nhận và được điều hòa bởi các biến đổi sinh lý và phân tử. Làm tổ là một quá trình gồm các bước chính như tiếp xúc, bám dính/gắn kết, xâm lấn và điều hòa miễn dịch. Các bước tuần tự này mang tính quyết định việc mang thai thành công. Hiểu được cơ chế sinh lý của giai đoạn đầu của thai kỳ sẽ giúp cải thiện đáng kể khả năng mang thai tự nhiên cũng như trong chu kỳ hỗ trợ sinh sản.
2.1 Điều kiện tử cung cho sự làm tổ
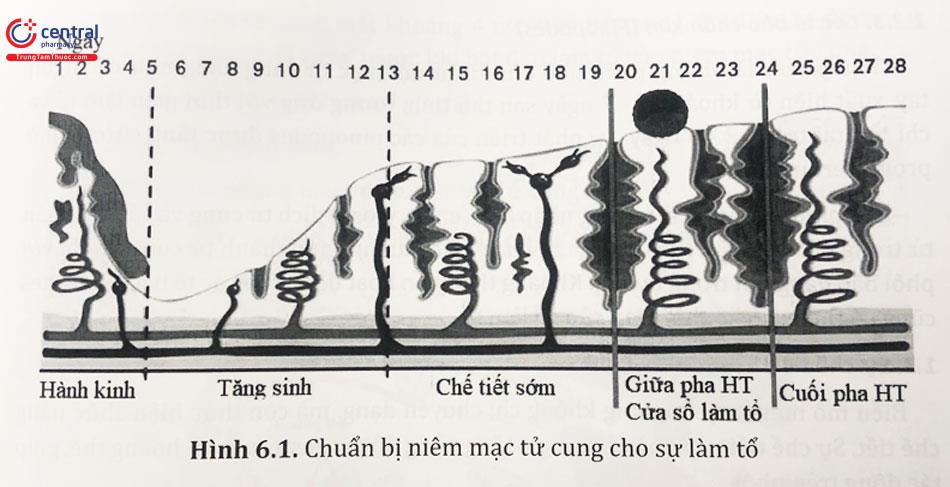
2.1.1 Cửa sổ làm tổ
Giai đoạn nội mạc tử cung sẵn sàng tiếp nhận phôi được gọi là “cửa sổ làm tổ” và kéo dài khoảng 3 - 4 ngày. Khoảng thời gian làm tổ xảy ra khoảng 6 ngày sau khi nồng độ hormone hoàng thể hóa (LH) đạt đỉnh và có thể kéo dài đến 9 ngày sau khi phóng noãn. Cửa sổ làm tổ được đặc trưng bởi những thay đổi các tế bào nội mạc tử cung, giúp hấp thụ chất lỏng trong tử cung. Những thay đổi này gồm sự biến đổi màng bào tương và đưa phôi nang đến gần nội mạc tử cung và cố định. Trong giai đoạn này, phối nang vẫn có thể bị đào thải ra ngoài buồng tử cung. Các hormone gồm estrogen và đặc biệt Progesterone giúp chuyển dạng chế tiết niêm mạc, là điều kiện “cửa sổ làm tổ”.
2.1.2 Sự chấp nhận của tử cung
2.1.2.1 Tiền màng rụng hóa
Nội mạc tử cung tăng sinh trở nên dày hơn, tái phân bố mạch máu và các tuyến cuộn xoắn, tăng tiết dịch. Những thay đổi này đạt cực đại khoảng 7 ngày sau khi phóng noãn. Bề mặt của nội mạc tử cung tạo ra loại tế bào tròn, được gọi là tế bào rụng (decidual cells), bao phủ toàn bộ phía buồng tử cung, xảy ra khoảng 9 - 10 ngày sau khi phóng noãn. Nếu không có thai, những tế bào này sẽ bong ra trong mỗi kỳ kinh nguyệt. Đồng thời, các tuyến tử cung giảm hoạt động và thoái hóa vào khoảng 8 - 9 ngày sau khi phóng noãn.
Các tế bào rụng bắt nguồn từ các tế bào mô đệm luôn có trong nội mạc tử cung. Tuy nhiên, các tế bào màng rụng tạo nên lớp màng mới - màng rụng (lớp decidua). Ngoài ra, phần còn lại của nội mạc tử cung thể hiện sự khác biệt giữa lớp mặt trong và phía đáy. Các tế bào viền tạo thành lớp đặc (zona compacta) của nội mạc tử cung, khác với lớp xốp (zona spongiosa) gồm các tế bào mô đệm khá xốp.
2.1.2.2 Màng rụng hóa
Quá trình màng rụng hoá sẽ diễn ra tiếp theo khi có thai làm tổ. Đây là một sự mở rộng của quá trình tiền màng rụng, phát triển thêm các tuyến tử cung, lớp đặc (zona compacta) và biểu mô của các tế bào rụng lót bên trong. Tế bào rụng chứa đầy lipid và glycogen và có hình dạng đa diện đặc trưng.
Phôi nang có thể đóng vai trò quan trọng trong kích hoạt sự phát triển và duy trì màng rụng hoá. Cơ sở của điều này là quá trình màng rụng hoá xảy ra ở mức độ cao hơn trong các chu kỳ có thụ thai. Phôi tiết ra các protease serine làm cho màng tế bào biểu mô khử cực và kích hoạt kênh Na* của biểu mô. Điều này kích hoạt dòng Ca và quá trình phosphoryl hóa CREB. Quá trình phosphoryl hóa CREB điều chỉnh sự biểu hiện của COX-2, dẫn đến giải phóng prostaglandin E2 (PGE2) từ các tế bào biểu mô. PGE2 hoạt động trên các tế bào mô đệm kích hoạt các con đường liên quan đến cAMP trong tế bào mô đệm dẫn đến quá trình màng rụng hóa.
Khi có sự làm tổ của thai, màng rụng gồm các phần:
- Màng rụng nền (Decidua basalis): là phần màng rụng nằm ở phía dưới đáy sau khi phôi làm tổ.
- Màng rụng bao (Decidua capsularis): phát triển quanh phôi thai, bao bọc nó vào nội mạc tử cung.
- Màng rụng thành (Decidua parietalis): Tất cả các phần còn lại trên bề mặt tử cung đều thuộc về màng rụng thành.
- Sau khi phôi làm tổ, màng rụng vẫn tiếp tục duy trì, đặc biệt trong quý đầu thai kỳ. Lớp đặc và lớp xốp vẫn quan sát được bên dưới màng rụng trong thai kỳ và các tuyến của lớp xốp tiếp tục chế tiết trong quý đầu.
2.1.2.3 Các tế bào chân kim (Pinopodes)
Pinopodes là những phần nhỏ nhô ra từ nội mạc tử cung dạng như các ngón tay, xuất hiện từ khoảng 5 - 7 ngày sau thụ tinh, tương ứng với thời gian làm tổ và chỉ tồn tại trong 2 - 3 ngày. Sự phát triển của các pinopodes được tăng cường nhờ progesterone.
Các pinopodes có chức năng nhập bào (endocytose) dịch tử cung và các đại phân tử trong đó. Nhờ vậy, thể tích của tử cung giảm xuống, giúp thành tử cung áp sát với phôi bào đang nổi trong buồng. Khoảng thời gian hoạt động của các tế bào pinocytes cũng có thể hạn chế thời gian cửa sổ làm tổ.
2.1.3 Sự chế tiết
Biểu mô niêm mạc tử cung không chỉ chuyển dạng mà còn thực hiện chức năng chế tiết. Sự chế tiết này nhờ vào sự gia tăng nồng độ progesterone từ hoàng thể, giúp tác động trên phôi.
Nguyên bào phôi nằm trong khoang tử cung khoảng 72 giờ trước khi làm tổ. Lúc này, nó chưa thể nhận được chất dinh dưỡng trực tiếp từ máu của mẹ, và phải nhờ vào chất dinh dưỡng được tiết vào khoang tử cung. Ngoài thành phần dinh dưỡng. nội mạc tử cung tiết ra một số protein phụ thuộc steroid, quan trọng cho sự phát triển và làm tổ, cholesterol và steroids. Quá trình làm tổ được tạo điều kiện thuận lợi hơn nữa nhờ sự tổng hợp các chất nền, các phân tử kết dính và các thụ thể bề mặt đối với các chất nền.

2.2 Điều kiện phôi trước làm tổ
2.2.1 Sự di chuyển của phôi sau thụ tinh
Sự thụ tinh thường xảy ra ở 1/3 ngoài vòi tử cung, ở vị trí đoạn bóng. Sau đó, phôi hình thành tiếp tục di chuyển trong vòi tử cung để đến buồng tử cung. Quá trình di chuyển trong vòi tử cung mất khoảng 4 ngày, và tiếp tục hiện diện tự do trong buồng tử cung thêm 2 - 3 ngày trước khi bắt đầu làm tổ vào niêm mạc tử cung.
Sự di chuyển của phôi nhờ vào 3 cơ chế:
- Nhu động của vòi tử cung.
- Hoạt động của nhung mao niêm mạc vòi tử cung.
- Luồng dịch chảy từ phía loa vòi tử cung vào buồng tử cung.
.jpg)
2.2.2 Điều kiện phôi trước làm tổ
Trên đường di chuyển, phôi phân bào rất nhanh, từ một tế bào ban đầu phân chia thành 2 rồi 4 tế bào bằng nhau, sau đó phân chia thành 8 tế bào và tạo nên phôi dâu, có hình dạng bên ngoài giống hình quả dâu. Sang ngày 5 phôi nang hình thành với các tế bào mầm nhỏ tạo thành lá nuôi có tác dụng nuôi dưỡng bào thai và các tế bào mầm to nằm ở giữa sẽ trở thành các lá thai, sẽ phát triển thành thai nhi.
Để có thể thực hiện quá trình làm tổ, điều kiện tiên quyết là phôi nang cần phải thoát ra khỏi màng thấu quang. Quá trình này được gọi là “phôi thoát màng”, được diễn ra nhờ vào các yếu tố của chính phôi nang. Nếu trứng không thụ tinh đặt vào trong buồng tử cung trong cùng điều kiện thì màng thấu quang vẫn nguyên vẹn. Đồng thời, yếu tố ly giải trong buồng tử cung cũng có vai trò quan trọng. Plasminogen, tiền chất của plasmin, được tìm thấy trong buồng tử cung, và các yếu tố từ phôi nang góp phần chuyển đổi thành plasmin hoạt động. Lý thuyết này đã được chứng minh bởi hiệu ứng gây ly giải in vitro của plasmin cũng như các thí nghiệm trên chuột.
2.3 Cơ chế làm tổ
2.3.1 Sự áp sát
Sự áp sát là hiện tượng tiếp xúc đầu tiên lỏng lẻo giữa phôi nang và nội mạc tử cung. Vị trí thường gặp trên nội mạc tử cung là các hốc nhỏ do khả năng tăng diện tích tiếp xúc với phôi nang đang có hình cầu. Đối với phôi nang, sau khi thoát màng thấu quang, có sự tiếp xúc trực tiếp giữa nguyên bào nuôi bên dưới và màng rụng của nội mạc tử cung. Gần như toàn bộ bề mặt của phôi nang có khả năng hình thành vị trí bám vào màng rụng.
Sự nhận diện các phân tử trên nguyên bào nuôi và biểu mô nội mạc tử cung làm trung gian cho sự tương tác ban đầu vẫn chưa hoàn toàn sáng tỏ. Tuy nhiên, protein được glycosyl hóa MUC1, một thành viên của họ Mucin đóng vai trò quan trọng. MUC1 là glycoprotein xuyên màng được biểu hiện ở đỉnh bề mặt của tế bào biểu mô nội mạc tử cung trong thời gian làm tổ ở người, làm trung gian cho sự gắn kết của phôi bào vào biểu mô tử cung.
Sự kết dính Sự kết dính là sự gắn chặt của phôi vào nội mạc tử cung sau khi áp sát. Các nguyên bào nuôi kết dính bằng cách xâm nhập vào nội mạc tử cung nhờ vào các vi nhung mao. Nguyên bào nuôi có các sợi liên kết, Collagen type IV và các integrins hỗ trợ quá trình kết dính này.
MUC16 là một chất xuyên màng được hiện diện ở bề mặt đỉnh của biểu mô tử cung. Chất mucin này ngăn phôi bào làm tổ vào vị trí không mong muốn trên biểu mô. Như vậy, MUC16 ức chế sự kết dính tế bào.
Giao tiếp giữa phôi và niêm mạc tử cung xảy ra ở giai đoạn này. Các tín hiệu truyền từ phôi nang đến nội mạc tử cung để tạo sự thích nghi tiếp theo. Chẳng hạn những thay đổi trong bộ khung tế bào của các tế bào rụng giúp tách các tế bào rụng khỏi liên kết với lớp đệm bên dưới, cho phép phôi bào xâm lấn sâu xuống dưới.
Thụ thể Proteoglycan: Một hệ thống thụ thể - ligand khác tham gia vào quá trình kết dính là các thụ thể proteoglycan, được tìm thấy trên bề mặt màng rụng của tử cung. Các đối tác của chúng, các proteoglycan, được tìm thấy xung quanh các nguyên bào nuôi của phôi nang. Hệ thống thụ thể - ligand này cũng hiện diện ở cửa sổ làm tổ.
2.3.2 Sự xâm nhập
Các phần nhô ra của các nguyên bào nuôi kết dính vào nội mạc tử cung tiếp tục tăng sinh và xâm nhập vào nội mạc tử cung, sau đó biệt hóa để thành một loại tế bào mới, gọi là hợp bào nuôi (syncytiotrophoblast). Phần còn lại của nguyên bào nuôi gọi là tế bào nuôi (cytotrophoblast). Hợp bào nuôi không được xác định là một loại tế bào, đúng hơn là một mô đa nhân.
Sự xâm lấn tiếp tục với các hợp bào nuôi chạm đến màng đáy bên dưới các tế bào rụng, xuyên qua nó và tiếp tục xâm nhập vào lớp đệm tử cung. Cuối cùng, toàn bộ phôi được chìm sâu vào nội mạc tử cung. Sau đó, các nguyên bào hợp bào tiếp xúc với máu mẹ và hình thành các nhung mao màng đệm. Đây là bước khởi đầu của việc hình thành nhau thai.

Sự xâm nhập của nguyên bào nuôi vào nội mạc tử cung được biểu hiện thông qua metalloproteinase MMP-2 và MMP-9. Hợp bào nuôi xâm nhập tử cung, tiếp cận nguồn cung cấp máu của mẹ, để thiết lập nền tảng cho tuần hoàn nuôi thai nhi.
Nguyên bào nuôi ngoài gai nhau (extravillous trophoblasts) là các tế bào từ nhung mao xâm nhập di chuyển vào cơ tử cung của mẹ. Những tế bào này tái cấu trúc các động mạch xoắn ốc để cải thiện và đảm bảo lưu lượng máu từ mẹ đến phôi thai đang phát triển. Quá trình này có thể xảy ra với các tĩnh mạch tử cung, ổn định để cải thiện tuần hoàn thai nhi và các chất thải chuyển hóa. Các nguyên bào nuôi cũng đã được ghi nhận là có thể di chuyển vào cơ thể mẹ và đã được tìm thấy trong các mô khác nhau.
2.4 Diễn biến chính quá trình làm tổ
2.4.1 Sự chế tiết
Các phôi bào tiết ra các yếu tố đa mục đích trong quá trình xâm nhập. Một số yếu tổ tự tiết, tự nhắm đích và kích thích chính nó xâm nhập sâu hơn vào nội mạc tử Các chất tiết làm lỏng liên kết giữa các tế bào rụng, ngăn cản sự đào thải phôi của cơ thể mẹ, thúc đẩy màng rụng hoa và ngăn hành kinh. Human Chorionic gonadotropin (hCG) là một yếu tố tăng trưởng tự tiết cho phôi nang. Yếu tố tăng trưởng giống Insulin 2 (IGF2) kích thích sự xâm lấn.
2.4.2 Sự tách rời tế bào
Các hợp bào nuôi tách rời các tế bào rụng bằng hai cách: thoái giáng các phân tử kết dính tế bào liên kết các tế bào rụng với nhau và thoái giáng chất nền ngoại bào.
Các phân tử kết dính tế bào bị thoái giáng do hợp bào nuôi tiết ra yếu tố hoại tử u-alpha (Tumour necrosis factor a-TNF-a), gây ức chế sự biểu hiện của cadherins và beta-catenin. Cadherins là các phân tử kết dính tế bào, và beta-catenin giúp gắn chúng vào màng tế bào. Do đó, liên kết giữa các tế bào rụng trở nên lỏng lẻo, cho phép các hợp bào nuôi và toàn bộ phôi xâm nhập vào nội mạc tử cung.
Chất nền ngoại bào bị phân hủy bởi serine endopeptidase và metalloproteinase. Các metalloproteinase như là collagenase, gelatinase và stromelysins gây tiêu hoá các type collagen và gelatin.
2.4.3 Ức chế miễn dịch
Phôi thai khác với các tế bào của người mẹ, và sẽ bị hệ thống miễn dịch của người mẹ đào thải nếu nó không tiết ra các chất ức chế miễn dịch. Một số tác nhân như yếu tố kích hoạt tiểu cầu (Platelet-activating factor), hCG, Prostaglandin E2, Interleukin 1-alpha, Interleukin 6, Interferon-alpha, yếu tố ức chế bạch cầu (leukemia inhibitory factor) và yếu tố kích thích khuẩn lạc (colony-stimulating factor).
Màng rụng hóa: các yếu tố từ phôi nang cũng kích hoạt sự hình thành cuối cùng của các tế bào rụng thành dạng phù hợp. Một số tế bào rụng ở gần phôi bào bị thoái hóa, cung cấp chất dinh dưỡng cho phôi.
2.4.4 Tắt kinh
HCG vừa hoạt động như một chất ức chế miễn dịch, vừa duy trì chức năng của hoàng thể, tiếp tục sản xuất các hormone sinh dục, giúp nội mạc tử cung tiếp tục phát triển, không hành kinh và là tín hiệu sớm nhất của thai kỳ.
2.4.5 Thất bại làm tổ
Sự thất bại trong quá trình làm tổ có thể liên quan đến sự tiếp nhận của tử cung (chiếm 2/3 trường hợp) hoặc do chính phôi thai. Khiếm khuyết trong khả năng tiếp nhận của tử cung có thể do tín hiệu bất thường của cytokine và nội tiết tố cũng như sự thay đổi thượng di truyền (của biểu mô nội mạc). Thất bại làm tổ liên tiếp có thể là một nguyên nhân gây vô sinh và cần được cải thiện bằng cách tối ưu hóa khả năng tiếp nhận của nội mạc tử cung. Đánh giá các marker sinh học làm tổ có thể giúp dự đoán kết quả mang thai và phát hiện các bất thường.
3 Tài liệu tham khảo
- PGS.TS.BS. Lê Minh Tâm và GS.TS.BS. Cao Ngọc Thành, ThS.BS. Trần Thị Như Quỳnh, ThS.BS. Nguyễn Đắc Nguyên, "Các vấn đề trọng yếu trong hỗ trợ sinh sản"
- Bruce M. Carlson (2019). Human Embryology and Developmental Biology. Sixth Edition. Elsevier.
- Cao Ngọc Thành, Lê Minh Tâm (2017). Nội tiết phụ khoa và Y học sinh sản. Nhà xuất bản Đại học Huế.
- Lê Minh Tâm, Nguyễn Thị Thái Thanh, Cao Ngọc Thành (2015). Nghiên cứu hiệu quả nuôi cấy phôi nang trong thụ tinh ống nghiệm. Tạp chí Phụ sản, tập 13 (01), 54-58. https://doi. org/ 10.46755/ vjog.2015.1.829.
- Linda M. Ernst, Eduardo D. Ruchelli Chrystalle Katte Carreon Dale S. Huff (2019). Atlas màu về mô học của thai nhi và trẻ sơ sinh. Springer Nature Thụy Sĩ AG.
- Minh Tam Le, Thai Thanh Thi Nguyen, Trung Van Nguyen, Hong Nhan Thi Dang, Quoc Huy Vu Nguyen (2021). Chuyển phôi nang sau khi nuôi cấy mở rộng phôi phân tách được bảo quản lạnh sẽ cải thiện kết quả của chu kỳ thụ tinh trong ống nghiệm. đông lạnh học. Tập 100, Trang 26-31. https:// doi.org/ 10.1016/ j.cryobiol.2021.04.003.
- Nguyễn Thị Thái Thanh, Nguyễn Văn Trung, Đặng Thị Hồng Nhạn, Lê Minh Tâm (2021). Nghiên cứu hiệu quả chuyển phôi nang từ phôi rã đông giai đoạn phân cắt. Tạp chí Phụ san, 19(1), 61-66. https://doi.org/10.46755/vjog.2021.1.1182
- Zhu, J. Y., Pang, Z. J., & Yu, Y. H (2012). Quy định về sự xâm lấn của trophoblast: vai trò của metallicoproteinase ma trận. Nhận xét về sản phụ khoa, 5(3-4), e137- e143
